 2016, Vol. 33
2016, Vol. 33文章信息
- 邢佳, 贺立娟, 王嘉麟, 王椿野, 郭蓉娟
- XING Jia, HE Li-juan, WANG Jia-lin, WANG Chun-ye, GUO Mao-juan
- 泻肝安神法对睡眠剥夺大鼠单胺类神经递质影响的研究
- Study of Xiegan Anshen method on monoamine neurotransmitters with rats after sleep deprivation
- 天津中医药, 2016, 33(3): 155-159
- Tianjin Journal of Traditional Chinese Medicine, 2016, 33(3): 155-159
- DOI: 10.11656/j.issn.1672-1519.2016.03.08
-
文章历史
- 收稿日期: 2015-11-26





现今社会生活节奏的加快使人们心理压力不断增加,失眠发病率逐年升高,已严重威胁到人们的生活质量及身心健康,中医认为压力过大、精神抑郁、情志不畅、肝气不舒、郁久化火、火热上扰心神,发为不寐,因此,不寐与肝、心两脏密切相关。很学者认同从肝论治失眠,认为肝在顽固性失眠辨证论治中起主导地位[1],失眠的核心病机为肝之刚柔不能相济[2]。同时,本课题组前期工作发现泻肝安神法标本兼顾对失眠患者的睡眠及情绪有很好的改善作用,临床取得较好疗效[3],但其疗效机制尚未阐明,以5-羟色胺(5-HT)为代表的单胺类神经递质,参与了睡眠的发生和维持,为本研究提供了思路。本研究旨在通过动物实验观察泻肝安神法治疗前后失眠大鼠脑干及海马单胺类神经递质含量测定,以探讨泻肝安神法干预失眠的疗效机制。具体实验方法和结果如下。
1 材料与方法 1.1 动物及分组健康雄性SPF级(SD)大鼠,体质量(250±20) g。由北京维通利华实验动物技术有限公司提供。大鼠适应环境1周后,用随机数字表法[4]分组为:空白组、失眠型组(模型组)、中药组、西药组。每组15只,共60只。
1.2 主要仪器及试剂5 -HT标准品Sigma,辛烷磺酸钠(1-OctanesulfonicAcid,Sodium Salt)色谱纯 J.T.Baker,乙二胺四乙酸(EDTA,Free acid)超纯 Amresco,甲醇(CH3OH)、 乙腈(CH3CN)色谱纯Fisher ChemAlert,0.5mol/L醋酸盐缓冲液(pH 6.0),5%四氢呋喃,邻苯二甲醛(OPA),双蒸馏水(DDW)
高效液相色谱仪(2695分离单元,2465 电导检测器,Waters),Empower色谱工作站。高速离心机(5417R,Eppendorf),旋涡混合器(MS2 Minishaker,IKA Works),超纯水器(Milli-Q Gradient A10,Millipore),标准型PH计(Basic PH Meter PB-20,Sartorius)。
1.3 实验药品地西泮:北京益民药业有限公司生产(国药准字H11020898),购自北京中医药大学东方医院。
中药汤剂:购自北京中医药大学东方医院中药房(柴胡10 g,郁金10 g,牡丹皮12 g,栀子10 g,当归12 g,白芍15 g,夜交藤15 g,酸枣仁20 g,川芎10 g,枳壳10 g,香附10 g,夏枯草15 g,黄芪10 g,茯苓15 g,刺五加15 g,炙甘草6 g),自行煎制。
按人体和大鼠比例公式计算药量。中药汤剂人体单次给药生药量为195 g,煎煮为250 mL中药汤剂,按公式计算折合大鼠给药量为17.5 g/kg,折合为具体药量平均为5.6 mL。西药片剂人体单次给药生药量1 mg,可溶解于10 mL纯净水中,按公式计算折合大鼠给药量为0.09 mg/kg,折合为具体药量平均为0.23 mL。
1.4 动物模型制备采用睡眠剥夺法造模,具体方法如下:将大鼠分别放入睡眠剥夺箱内的小平台上,自由进食、饮水,可进行转身、抬头、舔毛等活动,当其进入快速动眼相(REM)睡眠,由于全身肌张力降低、肌肉松弛,大鼠将落入平台下面的水中而惊醒,再重新爬上站台,继续在维持清醒状态下站立。反复剥夺睡眠7次,每次持续24 h(8∶00~8∶00),隔日1次,共14 d,以达到睡眠剥夺的效果。为了排除隔离、限制活动和水环境造成的应激影响,设立了大平台对照组,大鼠在大平台上可以睡眠而不会落入水中,其它环境与模型组相同,但大平台直径大,可保持完整的睡眠周期。正常对照组不接受睡眠剥夺。
1.5 组织取材及标本处理取材当天称质量后,10%水合氯醛(30 mg/kg)腹腔注射麻醉后直接断头取脑(整个过程应在5 min之内完成),从枕骨大孔向口腔侧进刀,剪开颅骨,剥去头顶骨,用镊子取出整个脑组织,冰盘上分离出海马、脑干及下丘脑,冰冻生理盐水漂洗后滤纸拭干表面水分,电子天平称质量,标记称质量后分别装入1.5 mL Eppendorf管,放置于 -80 ℃冰箱保存,待样品集齐后成批检测。检测前将称质量后的脑组织(20~50 mg)。放入预冷至4 ℃的0.4 mol/L高氯酸90 mg中,冰浴中匀浆2~3 min,并沉淀30 min去除蛋白。于冰浴下快速玻璃匀浆,静置10 min,4 ℃条件下14 000×g低温离心15 min,取上清液,用0.22 μm滤膜过滤,-80 ℃保存待测[氨基酸检测用Costar(Spin-X,0.22 μm)离心式过滤器7 000 r/min离心过滤,再用0.05 mol/L 高氯酸10倍稀释,取20 μL放入自动进样器中待测]。
1.6 色谱条件预柱:SHISEIDO Guard Cartridge,Capcell C18 MG S-5;色谱柱:ESA MD-150(3.2 mm×150 mm);流动相:90 mmol/L 磷酸二氢钠,50 mmol/L 柠檬酸,1.7 mmol/L 1-辛烷磺酸钠,50 μmol/L EDTA,10% 乙腈,pH 约3.00。流速:0.6 mL/min;进样量:20 μL;柱温:30 ℃;设置4道电势为:-150、200、350、500 mV。
1.7 统计处理将有关数据进行数字化处理,输入计算机,采用Microsoft Office Access 2003建立数据库,SPSS 17.0统计软件包进行分析,计量资料以均数±标准差(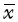 ±s)表示。符合正态分布的计量资料组间比较采用单因素方差分析,两两比较时若方差齐采用snk-q检验,若方差不齐采用Dunnett’s T3检验;偏态分布的计量资料,组间比较采用Kruskal-Wallis秩和检验,组间两两比较采用Nemenyi检验。P<0.05为差异有统计学意义。
±s)表示。符合正态分布的计量资料组间比较采用单因素方差分析,两两比较时若方差齐采用snk-q检验,若方差不齐采用Dunnett’s T3检验;偏态分布的计量资料,组间比较采用Kruskal-Wallis秩和检验,组间两两比较采用Nemenyi检验。P<0.05为差异有统计学意义。
实验结果统计提示睡眠剥夺后,模型组大鼠脑干区的5-HT含量较空白组明显下降,海马区也相对降低,但变化无统计学差异。而在泻肝安神汤干预1周之后,中药组在脑干区及海马区的5-HT含量均显著高于模型组,且在海马区高于西药组。见表1。
±s)
Tab.1 Contents of 5-HT in brainstemand hippocampus
with rats of each groups(±s)
| μg/L | ||||
| 组别 | 动物数 | 脑干 | 左侧海马 | 右侧海马 |
| 空白组 | 15 | 513.46±104.93 | 189.47±106.21 | 110.45±044.93 |
| 模型组 | 15 | 304.02±133.36# | 101.42±045.17 | 085.05±055.06 |
| 西药组 | 15 | 424.52±116.54 | 281.21±140.26* | 212.23±062.58 |
| 中药组 | 15 | 476.72±191.57* | 297.34±189.41* | 528.64±358.49#* |
| 注:与模型组比较,*P<0.05;与空白组比较,#P<0.05。 | ||||
实验结果统计提示睡眠剥夺后,模型组大鼠脑干区的5-HIAA含量较空白对照组明显下降,海马区也相对降低,但变化无统计学差异。而在灌胃给药1周之后,中药组和西药组均在脑干区及海马区的5-HIAA含量均显著高于模型组,并且高于空白对照组。见表2。
±s)
Tab.2 Contents of 5-HIAA in brainstemand hippocampus
with rats of each groups(±s)
| μg/L | ||||
| 组别 | 动物数 | 脑干 | 左侧海马 | 右侧海马 |
| 空白组 | 15 | 447.22±060.10 | 219.84±091.43 | 268.20±0160.48 |
| 模型组 | 15 | 279.82±131.16# | 153.58±053.02 | 233.77±0102.65 |
| 西药组 | 15 | 533.94±220.41* | 198.58±081.97 | 1 480.41±0509.29#* |
| 中药组 | 15 | 508.89±148.99* | 262.32±130.61 | 2 213.10±1155.86#* |
| 注:与模型组比较,*P<0.05;与空白组比较,#P<0.05。 | ||||
实验结果统计提示睡眠剥夺后,模型组大鼠脑干区的DA含量较空白对照组明显升高,海马区左右各有差异,但均无统计学差异。而在灌胃给药1周之后,中药组较模型组在脑干区DA递质含量减低,在海马区反而升高。西药组在脑干区及右侧海马较模型组DA递质含量无明显变化,在左侧海马明显升高。见表3。
±s)
Tab.3 Contents of DA in brainstemand hippocampus with
rats of each groups(±s)
| μg/L | ||||
| 组别 | 动物数 | 脑干 | 左侧海马 | 右侧海马 |
| 空白组 | 15 | 035.30±22.21 | 183.22±090.47 | 045.49±24.91 |
| 模型组 | 15 | 100.26±21.85# | 090.14±051.63 | 086.88±56.09 |
| 西药组 | 15 | 118.19±39.01# | 178.49±070.69* | 030.50±10.20 |
| 中药组 | 15 | 067.52±18.11#* | 223.63±153.50* | 109.86±42.05#* |
| 注:与模型组比较,*P<0.05;与空白组比较,#P<0.05。 | ||||
实验结果统计,西药组在脑干区的DOPAC含量高于空白对照组。其余各组间在各不同脑区DOPAC递质含量变化无统计学差异。见表4。
±s)
Tab.4 Contents of DOPAC in brainstemand hippocampus
with rats of each groups(±s)
| μg/L | ||||
| 组别 | 动物数 | 脑干 | 左侧海马 | 右侧海马 |
| 空白组 | 15 | 12.86±06.45 | 47.35±20.51 | 36.32±29.20 |
| 模型组 | 15 | 26.69±08.58 | 29.88±12.30 | 13.82±14.18 |
| 西药组 | 15 | 33.16±19.70* | 40.99±25.41 | 86.90±71.93 |
| 中药组 | 15 | 26.03±07.48 | 42.41±22.47 | 66.03±30.01 |
| 注:与模型组比较,*P<0.05;与空白组比较,#P<0.05。 | ||||
实验结果统计提示睡眠剥夺后,模型组大鼠HVA递质含量在脑干区较空白对照组升高,在海马区降低。而在灌胃给药1周之后,中药组HVA递质含量较模型组在海马区升高,在脑干区无明显统计学差异。西药组HVA递质含量较模型组在右侧海马区升高,在脑干和左侧海马无明显统计学差异。见表5。
±s)
Tab.5 Contents of HVA in brainstemand hippocampus
with rats of each groups(±s)
| μg/L | ||||
| 组别 | 动物数 | 脑干 | 左侧海马 | 右侧海马 |
| 空白组 | 15 | 16.44±07.2 | 20.49±1000 | 016.46±013.10 |
| 模型组 | 15 | 29.26±07.29# | 10.42±4.43# | 010.85±161.87 |
| 西药组 | 15 | 29.31±11.27# | 09.86±3.48# | 101.18±051.59#* |
| 中药组 | 15 | 30.51±09.92# | 16.92±7.67* | 082.73±072.18#* |
| 注:与模型组比较,*P<0.05;与空白组比较,#P<0.05。 | ||||
实验结果统计提示睡眠剥夺后,模型组大鼠NE含量在脑干及右侧海马较空白对照组升高,在左侧海马区也有升高趋势,但无统计学差异。而在灌胃给药1周之后,中药组和西药组均在右侧海马NE含量较模型组降低,在脑干及左侧海马含量变化无明显统计学差异。见表6。
±s)
Tab.6 Contents of NE in brainstemand hippocampus with
rats of each groups(±s)
| μg/L | ||||
| 组别 | 动物数 | 脑干 | 左侧海马 | 右侧海马 |
| 空白组 | 15 | 0648.40±237.57 | 1 974.18±982.91 | 254.97±103.91 |
| 模型组 | 15 | 0867.75±209.75# | 2 193.95±1 245.37 | 875.36±523.78# |
| 西药组 | 15 | 1015.48±255.82# | 1 887.17±659.94 | 514.87±191.31* |
| 中药组 | 15 | 0827.65±139.66 | 1 263.15±49.25 | 158.95±079.11* |
| 注:与模型组比较,*P<0.05;与空白组比较,#P<0.05。 | ||||
综上结果表明,大鼠剥夺睡眠后脑干单胺类神经递质5-HIAA、5-HT明显下降,DA、NE、HVA明显升高,DOPAC无明显变化。而在海马区域各单胺类神经递质含量变化不显著,且左右略有差异,仅HVA含量在左侧海马有所降低,NE含量在右侧海马升高。采取泻肝安神法治疗后大鼠脑干5-HT、5-HIAA含量显著升高,DA含量明显下降;此外DOPAC、NE有下降趋势,但无统计学差异。而在双侧海马5-HT、5-HIAA、DA、HVA升高,NE下降。采用地西泮片治疗后大鼠在海马区的单胺类神经递质变化与中药组结果一致,但在脑干仅有5-HIAA升高,而5-HT、DA则没有明显变化。
3 结论与讨论 3.1 大鼠剥夺睡眠造模后神经递质的变化研究发现剥夺睡眠能导致动物在探究活动、自发活动[5]、焦虑行为上发生异常变化[6, 7],同时对大鼠海马结构的蛋白表达有一定影响[8]。近年来多项研究表明[9, 10, 11, 12, 13, 14],剥夺睡眠不仅后脑内5-HT、5-HIAA下降,DA、NE升高,给予药物治疗后这些递质变化被纠正。但血浆内的神经递质变化结果并不一致,有研究认为,剥夺睡眠后血浆内NE、5-HT升高,DA下降[15],不同的造模方法对血浆内神经递质含量的变化影响是不同的[16]。但是在脑组织的单胺类神经递质含量测定方面,无论是水平台法的物理方法还是腹腔注射对氯苯丙氨酸(PCPA)的化学方法建立的失眠模型,均得出同样的结果,这可能提示脑内的单胺类神经递质含量变化相对稳定。本实验采用水平台法剥夺睡眠建立失眠模型,实验结果提示大鼠剥夺睡眠后脑干单胺类神经递质5-HIAA、5-HT明显下降,DA、NE、HVA明显升高,DOPAC无明显变化。该结果表示剥夺睡眠后大鼠神经递质发生改变,这与目前对于失眠机制及动物模型建立的各项研究结果相符,证明造模成功有效。而在海马区域各单胺类神经递质含量变化不显著,且左右略有差异,但在双侧海马5-HT、5-HIAA含量均有下降趋势,NE有升高趋势,与脑干区的单胺类递质含量变化趋势呈现一致,但无统计学差异。考虑原因:一是可能由于海马体积小,单胺类递质含量较少,并且实验采取两侧分别测量,样本量不大,故测量值不易出现明显差异;二是海马中虽然含有许多与睡眠调节相关的神经递质[17, 18],但其与中缝核、杏仁核、蓝斑、丘脑、下丘脑等调节睡眠的神经结构间有着复杂的神经联系[19],它与特定的学习和记忆有关,参与了记忆的获取,以及在别的结构建立持久和可追溯的记忆。海马功能主要与认知记忆相关,本研究主要为剥夺睡眠,观察药物改善睡眠的疗效机制,造模及给药时程均较短,故本研究未发现海马区的单胺类递质含量的明显变化。
3.2 泻肝安神法对大鼠脑内单胺类递质的影响睡眠是一个重要的生物学过程,通过多个脑区和神经调节物质交互错杂作用来调节的。涉及到中枢神经系统不同层次众多的神经网络和一系列的神经递质、神经调节物质[20]。5-HT、NE、DA等多种神经递质均参与睡眠与觉醒的调控[21, 22]。失眠是一种神经节律失常性疾病,目前认为单胺类递质参与睡眠的生理调节过程,这几种神经递质的交互作用共同维持觉醒和睡眠及非快速动眼睡眠期(NREM)和REM睡眠的周期变化[15]。同时5-HT受体又与DA、NE和神经内分泌系统有着相互调节的作用[23]。而目前对于失眠的有效治疗均是通过调整其神经递质含量而实现的。本研究发现泻肝安神法可升高脑干内的5-HT和5-HIAA含量,降低脑干内DA含量;同时升高海马的5-HT、5-HIAA、DA、HVA含量,降低海马内NE含量。采用地西泮片治疗后大鼠在海马区的单胺类神经递质变化与中药组结果一致,但在脑干仅有5-HIAA升高,而5-HT、DA则没有明显变化。说明泻肝安神法与地西泮均可调节睡眠剥夺大鼠的神经递质,提示其疗效机制可能与改变不同部位脑内神经递质的含量有关。且泻肝安神法对于单胺类神经递质的影响靶点要多于地西泮片,其治疗失眠的疗效机制可能与综合的调节单胺类神经递质水平,纠正其紊乱相关。
| [1] | 姜 华.顽固性失眠从肝论治临证探讨[J].天津中医药,2005, 22(4):316-317. |
| [2] | 赵志付,吴志奎,王彩凤,等.胆胀病的中医刚柔辨证研究概要[J].中国医药导刊,2011,13(9):1590. |
| [3] | 邢 佳,王嘉麟,王椿野,等,疏肝泻火养血安神法干预失眠症的临床研究[J].环球中医药,2013,6(7):500-504. |
| [4] | 苗明三.实验动物和动物实验技术[M].北京:中国中医药出版社,1997:142. |
| [5] | Prut L Belrung C.The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors review[J]. Eur J Pharmacol,2003,463(1-3):3-33. |
| [6] | Pellow S,Chopin P,File S,et a1.Validation of open:closed arm entries in an elevated plus-maze as a meagure of anxiety in the rat[J].Neurosci Meth,1985,14:149-167. |
| [7] | 范新六,李 峰,宋月晗,等.四慧合剂对睡眠剥夺大鼠旷场及高架十字迷宫行为的影响[J].天津中医药,2011, 28(2):142-144. |
| [8] | 王秀云,李积胜,刘公望,等. 睡眠剥夺对亚健康形成的影响及相关机制[J].天津中医药,2006, 23(2):108-111. |
| [9] | 周春权,曾雪爱,黄俊山. 中药松郁安神方对失眠大鼠血浆中单胺类神经递质的影响[J].中华中医药学刊,2012,30(6):1323-1324. |
| [10] | 刁远明,陈 群,陈新林.安寐汤对失眠大鼠脑组织单胺类神经递质的影响[J].广州中医药大学学报,2011,28(4):409. |
| [11] | 刁远明,陈 群.安寐汤对失眠大鼠脑组织多巴胺含量的影响[J].中医中药导报,2011, 8(17):105. |
| [12] | 刘 彬,臧佩林.保神开郁冲剂影响失眠鼠中枢神经递质的实验研究[J].辽宁中医杂志,2000, 27(2):92. |
| [13] | 兰 妮.失眠大鼠电针后不同时相单胺类和氨基酸类递质的变化[D]. 优秀硕士毕业论文库,2010,6. |
| [14] | 滕 晶.醒寤晨方与安寐晚方协同用药对失眠大鼠脑内单胺类神经递质的影响[J].天津中医药,2007, 24(1):51-53. |
| [15] | 钟静喻. 睡眠剥夺对大鼠血浆NE、DA、5-HT及认知功能的影响[D].优秀硕士毕业论文库,2010,6. |
| [16] | 王雅丽. 睡眠剥夺对大鼠学习激励和神经递质的影响及中西药敢于作用[C].优秀硕士毕业论文库,2011:3. |
| [17] | A mat J,Matus-Amat P,Watkins LR,et al.E scapable and in escapable stress differentially and selectively alter extracellular leves of 5-HT in the hippocampus and dorsal periaqueductal gray of the rat[J].Brain Res,1998,797(1):12-22. |
| [18] | Loy R,Kozieill DA,Lindsey JD.Noradrenergic intervation of the adult rat hippocampal formation[J]. Comp Neurol,1980,189:699-710. |
| [19] | 张培林.神经解剖学[M].北京:人民卫生出版社,1999:477-492. |
| [20] | Krueger JM,0bal FJ,Kapas L,et a1.Brain organization and sleep function[J].Brain Res,1995,69(2):177-185. |
| [21] | Maloney KJ,Mainville L,Jones BE.Diffferential c-Fos expression in cholinergic,mono—aminergic,and GABA ergic cell groups of the pontomesencephalic tegmentum after paradoxical sleep deprivation and recovery[J].J Neurosci, 1999,19(8):3057-3072. |
| [22] | Youngblood BD,smagin GN,Elkins PD.The effects of paradoxical sIeep deprivation and valine on spatial learing and brain 5-HT metabo1ism[J].Physiol Behav,1999,67(5):643-649. |
| [23] | 张 雪.酸枣仁的化学成分和药理作用研究进展[J].食品工业技术,2009,30(3):348. |