 2026, Vol. 43
2026, Vol. 43文章信息
- 何雪, 孙一珂, 刘雷蕾.
- HE Xue, SUN Yike, LIU Leilei.
- 基于长夏溃疡结肠炎的发病机制探讨“脾应长夏”的生物学内涵
- Discussion on the biological connotation of "spleen corresponding to late summer" based on the pathogenesis of ulcerative colitis in late summer
- 天津中医药, 2026, 43(1): 62-66
- Tianjin Journal of Traditional Chinese Medicine, 2026, 43(1): 62-66
- http://dx.doi.org/10.11656/j.issn.1672-1519.2026.01.11
-
文章历史
- 收稿日期: 2025-08-15

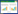



随着阴阳学派和五行学派的深入交融,“五时五脏”不仅成为了阴阳学派五行化发展的成熟理论,也在《黄帝内经》的时脏配属体系中占据了主流地位[1]。“脾应长夏”作为“五时五脏”的重要组成部分,不仅是中医“藏象学说”的重要思想,更是融合了“天人相应”的中医特色理论。长夏一般指农历六月,处于夏秋相交之际,是一年中最潮湿、最炎热的季节。作为一种慢性非特性肠道炎症,溃疡性结肠炎(UC)的典型临床表现为腹泻、黏液脓血便,同时可伴有腹痛、里急后重、发热等全身症状[2]。UC易于长夏发病[3-5]可能与人体“脾应长夏”的生理特性与长夏湿热弥漫的气候特征有关。然而目前关于“脾应长夏”科学内涵以及气候湿温变化对于UC发病机制影响的相关研究,尚属空白。
笔者团队通过前期研究发现:UC易于长夏发病,中医病机与“脾应长夏”的生理特性密切相关,肠道菌群与中医“脾”关系密切,其结构组成可受到季节气候的影响而发生相应的变化,可作为脾与长夏相通应的中介物质。病理情况下,肠道菌群的结构组成在长夏发生异常,紊乱的肠道菌群可通过活性氧(ROS)/缺氧诱导因子-1α(HIF-1α)途径介导经典活化型巨噬细胞(M1)/替代活化型巨噬细胞(M2)的极化失衡,进而诱发UC。笔者将“脾应长夏”理论与季节气候变化、肠道微生态、现代免疫学成果进行多维整合,旨在揭示“脾应长夏”的生物学本质,阐明“天人相应”的科学内涵,并围绕“肠道菌群-ROS/HIF-1α-巨噬细胞M1/M2极化”这一调节链的失衡,为防治UC的季节性发病提供新的干预靶点。
1 UC易于长夏发病长夏一般指农历六月,长夏处于夏、秋两季之间,涵盖了小暑、大暑、立秋、处暑4个节气,现代研究发现UC易于长夏发病:如1项纳入409例UC患者的关于UC发作月份的季节性研究报告显示:在可统计的1 030次症状中,8月出现发作高峰[3]。王嘉源等[5]对192例UC患者进行回顾性分析发现患者入院时间存在季节性规律,以8月份入院例数最多。根据UC腹泻、黏液脓血便、腹痛等典型临床表现,中医将UC归为“肠澼”“痢疾”“泄泻”等研究范畴。在中医古籍中屡有“痢疾”“肠澼”“泄泻”等在长夏多发的记载:如《素问·金匮真言论篇》中言:“长夏善病洞泄寒中”。《素问·至真要大论》曾记载:“炎暑至……腹满痛溏泄”“暴热消烁……少腹痛,下沃赤白。”李东垣也曾在《东垣十书》中言:“今时值长夏,湿热大盛,正当客气胜而主气弱也,故肠之病甚。”《医宗必读》中记载:“夫痢起夏秋,湿蒸热郁,本乎天也。”由此可见,UC具有易于长夏发病的季节性发病规律。
2 基于“脾应长夏”探讨UC长夏发病 2.1 季节气候变化对人体生理病理产生的重要影响“天人相应”是作为一种人与宇宙的协同共振关系,其以气机的生、长、化、收、藏为核心,以阴阳矛盾运动为动力,以五行生克制化为自稳调节器[6]。四时更迭,气候递嬗:春气温和、夏令炎热、长夏湿重、秋来凉爽、冬时严寒。人之生理亦随之外显“生、长、化、收、藏”之节律。《灵枢·五窿津液别》曾述“天暑衣厚则腠理开,故汗出……天寒则腠理闭,气温不行,水下留于膀胱,则为溺与气”,正是机体应天变动的表现,人体排泄汗液与尿液的功能受到季节气候变化的影响而表现出:冬季多尿少汗,夏季多汗少尿的适应性调节。由此可见,人体的生理功能可根据自然界季节气候的更替而发生相应的适应性调节,这是机体顺应自然的一种调节机制。季节气候除了对人体的生理功能产生相应的影响,对人体的病理也会产生重要的影响。
综上所述,人体存在着人与自然界共振的调节机制,自然界季节气候的变化对人体病理生理都有着重要的影响。从季节气候变化的角度出发研究其对人体的影响,在生理、病理两个层面都有丰富的文献记载作为研究支持,并且符合中医学“天人相应”理论与“五脏应时”理论的认识,因而结合季节气候节律性变化阐发藏象系统生理、病理变化的生物学机制具有可行性。
2.2 正确把握“脾应长夏”的理论实质目前关于“脾应长夏”的理论实质存在多种认识,如“脾不独主于长夏之时”“脾主长夏”“脾虽旺于四时,而以长夏为最旺”“脾旺于长夏,长夏季节多见脾病变”等[7]。笔者团队认为“脾应长夏”的理论实质为:脾是机体在长夏季节自稳调节系统。脾与胃相表里,两者一升一降,共同构成了消化系统的轴心,脾升胃降的生理特性概括了整个消化系统的功能。脾喜燥恶湿,主运化、升清;胃喜润恶燥,主受纳、降浊。两者纳运相合、升降相因、燥湿互济,共同维持“应长夏而变”的动态平衡。然而长夏湿气当令,外湿最易困脾,导致运化、升清之力相对减弱;湿与热合,反助胃之受纳、通降功能相对亢进,于是出现“脾弱胃强”的阶段性特征。此时,脾胃系统借此失衡—再平衡机制,主导并协调其余四脏,完成季节转换的整体调节;而在其他季节,脾胃则处于从属地位,通过其长养转化之力,随四时进退,协助其他四脏维持节律性稳态。
2.3 “脾应长夏”是UC长夏发病的核心病机UC的病位虽然在大肠,但其发病与脾胃密切相关。由于长夏季节脾虚胃强的功能状态处于支配地位,因而病理情况下,正如《素问·阴阳应象大论》所云:“清气在下,则生飨泄;浊气在上,则生瞋胀”,脾主运化功能减弱,脾失健运,饮食物和津液难以吸收转运,水谷之精微与津液不能向上向外扩散,也不能上传于肺。同时,脾升不足,胃降有余,脾胃升清降浊功能紊乱,清者不升,浊者独降,清浊相干,顿发为腹泻、腹痛、腹胀等UC的典型临床表现。此外,长夏湿热,若与暑结,气血搏结并裹挟肠中秽浊,则会出现下痢脓血、肛门灼热、里急后重等临床表现。
由此可见,长夏主湿,脾作为机体在长夏季节发挥主要调节作用的自稳调节系统,其与长夏相适应的生理特性以及长夏季节湿热弥漫的气候特征,与UC长夏发病存在紧密的联系。而在现代医学的研究中,UC的发病机制中涉及免疫功能的异常,而机体的免疫应答水平受温度和昼夜节律等因素的调节,可以呈现季节性改变[6-7]。长夏季节位于夏秋之间,是一年中气候最潮湿、最炎热的季节,UC在长夏发病,其发病机制可能与长夏季节湿温变化介导的异常免疫有关。
3 巨噬细胞M1/M2极化失衡是UC发病的核心环节作为先天固有免疫的重要组成成分,巨噬细胞在机体的炎症和宿主防御中发挥着核心作用。其功能表型具有高度可塑性,在各种因素(如微生物产物、受损细胞、活化的淋巴细胞等)或在不同的病理生理条件的诱导下,巨噬细胞可极化为不同的功能表型,即巨噬细胞M1型和M2型[8]。
在UC发生发展的过程中,M1型巨噬细胞虽可吞噬病原体和微生物,并对炎症消退和促进伤口愈合发挥关键作用,但其同步分泌的大量促炎细胞因子[如肿瘤坏死因子-α(TNF-α)、白细胞介素-1β(IL-1β)等]以及高水平的诱导型一氧化氮合酶(iNOS)和活性氧(ROS),反而放大炎症级联,并延缓组织损伤的修复和伤口的愈合[9]。相反,M2型巨噬细胞通过分泌TGF-β1、白细胞介素-10(IL-10)等抗炎细胞因子,介导碎片清除、血管新生及细胞外基质重塑,从而减轻肠道损伤[10]。肠道免疫稳态的维持依赖于M1与M2亚群的动态平衡,而巨噬细胞M1/M2的极化失衡通常发生在各种炎症性疾病之中,越来越多的研究表明巨噬细胞M1/M2极化失衡是UC发病的关键环节,可作为治疗UC的潜在靶点,这也使得通过调控巨噬细胞的极化表型来控制肠道炎症和恢复组织损伤成为了一种治疗UC的新兴手段[11-12]。
4 肠道菌群对巨噬细胞M1/M2极化平衡的调控作用为“脾应长夏”本质内涵的研究提供新思路肠道菌群是一个存在于宿主消化道内的由细菌、古细菌、真菌和病毒等微生物构成的复杂微生态系统。研究表明,长夏病理情况下,肠道菌群的结构组成发生紊乱,表现为以拟杆菌门为主,F/B值下降,紊乱的肠道菌群可通过ROS/HIF-1α途径调控巨噬细胞M1 /M2的极化平衡进而诱导UC的发生。
4.1 肠道菌群与脾关系密切中医里脾为后天之本,气血生化之源。正如《素问·经脉别论》所言:“饮入于胃,游溢精气,上输于脾,脾气散精”,作为脾最主要的生理功能,脾主运化指的是脾具有将饮食水谷化为精微,以及将精微物质吸收并转输全身的作用。中医对水液代谢的异常也多责之于脾,《素问·至真要大论篇》有云:“诸湿肿满,皆属于脾。”由此可见,脾在人体饮食物的消化、吸收、水液的代谢方面发挥着重要的作用。在现代研究中,肠道微生态广泛介入机体营养物质的消化、吸收和合成以及水液代谢等生理过程,如分解碳水化合物、蛋白质、脂类,合成多种维生素、胆汁酸、有益的短链脂肪酸,以及提高铁等金属离子的吸收率等等[13]。从这个角度看,肠道菌群与脾关系密切,可归属于中医“脾”的研究范畴。脾的生理功能正常与否,直接决定人体正气是否充沛,构成机体抵御外邪入侵的关键屏障。正气指机体的抗病能力,包括气血津液等有形物质和脏腑经络功能等在内的机体功能状态。而肠道菌群及其代谢产物可通过为机体提供能量、加强肠道黏膜屏障的功能、增强免疫和抵御病原微生物入侵,从而维持肠道健康,因此肠道菌群又被称为“人体的第二大免疫系统”[14]。肠黏膜屏障功能保障了胃肠道正常的消化吸收功能[15],与机体的免疫功能都可作为中医“正气”的生物学基础。因此,从这个角度出发,同样可以论证肠道菌群与脾存在着一定的相似性。同时,现代学者在脾虚模型大鼠肠道菌群分布及时效性研究中发现:与对照组大鼠肠道菌群物种丰富度及显著差异物种数相比,脾虚组存在明显差异[16]。具有健脾益气作用的代表方剂四君子汤能有效扶植UC大鼠正常菌群的生长,改善和调节肠道微生态的分布[17]。这些研究都从病理角度反证了肠道菌群与脾之间的密切联系。
综上所述,肠道菌群参与人体对饮食物的消化和吸收、水液代谢、免疫等生理功能,无论是从生理角度出发,还是从病理角度反证,肠道菌群与脾都存在一定的相似性与同一性,两者关系密切,肠道菌群可归属于中医学“脾”的研究范畴。
4.2 肠道菌群可作为脾与长夏相通应的中介物质肠道菌群是一个极其复杂的微生态系统,其变化受到多种因素的影响[18]。近年的研究发现肠道菌群与调控昼夜节律的时钟基因存在双向调控关系,肠道菌群的变化不仅存在昼夜节律[19],还受到季节气候变化的影响。有学者[20]联合16S rDNA扩增子测序与靶向代谢组学解析长夏肠道微生态的季节图谱,结果显示:生理状态下,该时段菌群以厚壁菌门占主导,属水平则以乳杆菌属丰度最高。另有研究[21]利用基因测序技术观察季节更替对大鼠肠道菌群的影响,结果显示:生理条件下,夏季与冬季大鼠在空肠、回肠末端及升结肠的微生物群落结构显著不同,夏季以厚壁菌门为优势菌群,F/B比值升高。而在病理情况下,研究[22]基于“脾恶湿”理论观察长夏湿困证模型发现:病理状态下,厚壁菌门乳酸菌丰度显著下降。该类益生菌原本通过拮抗致病菌、促进共生菌增殖及调控物质能量代谢维系微生态稳态并改善胃肠功能[23],其优势丧失可能是湿邪困脾的重要微生态标志。肠道菌群的结构受到季节气候变化的影响,进而表现出一定的季节性特征。肠道菌群与“脾”关系密切,可归属于“脾”的研究范畴,基于肠道菌群在长夏的季节性变化以及肠道菌群与脾之间的密切联系,笔者团队提出肠道菌群可作为脾与长夏相通应的中介物质,以肠道菌群-免疫互作为切入点,揭示“脾应长夏”的生物学内涵具备可行性。
4.3 肠道菌群可通过ROS/HIF-1α途径介导M1/M2极化平衡UC的发病机制涉及肠道菌群介导的异常免疫。研究表明,肠道中脆弱拟杆菌等革兰氏阴性菌(拟杆菌门)可以通过降低线粒体能量代谢减少腺嘌呤核苷三磷酸(ATP)合成[24]。在低供能情况下,巨噬细胞更易分化成M1型巨噬细胞[25],并分泌促炎因子TNF-α、IL-1β等。M1型巨噬细胞和M2型巨噬细胞在主要的能量供应途径上存在显著差异,其线粒体呼吸链的适应在抗菌宿主防御中发挥了重要作用。研究显示,来源于线粒体内膜的呼吸链底物端的活性氧(ROS)在巨噬细胞极化的调控中起着重要的作用,而肠道微生物可以通过线粒体呼吸链来调控ROS的产生[26]。肠道菌群会释放病原体相关分子模式(PAMPs),通过激活PAMPs识别受体(PRR),靶向作用于线粒体。位于质膜和内质网膜上的Toll样受体(TLR)是识别微生物并引起免疫反应的一组蛋白质识别受体,其可识别细菌、病毒、真菌及寄生虫来源的PAMPs。细菌特异性抗原脂多糖(LPS)是一种源自革兰氏阴性细菌的促炎分子,拟杆菌门贡献了肠道中LPS产量的绝大部分[27]。LPS处理会通过TLR4途径作用于巨噬细胞中线粒体,降低细胞线粒体呼吸链复合体Ⅰ-Ⅴ酶活性和线粒体呼吸链的功能,致使ROS生成增加,导致线粒体氧化磷酸化(OXPHOS)功能降低[24],诱导巨噬细胞向M1型极化。同时,ROS可与NO产生协同效应,激活HIF-1α转录因子活性[28]。HIF-1α是细胞适应缺氧的主要调节因子,也能够诱导巨噬细胞向M1型极化。
研究表明,长夏病理情况下,肠道菌群结构发生明显改变,以拟杆菌门为主,F/B值明显降低[20]。因而长夏病理情况下,肠道菌群对巨噬细胞M1/M2极化调控以拟杆菌门发挥的效应为主,即主要促进巨噬细胞向M1型极化,从而介导了巨噬细胞M1/ M2极化失衡,诱导了UC的发生。见图 1。

|
| 注:PAMPs,原体相关分子模式;PRR,PAMPs识别受体;ROS,活性氧;HIF-1α,缺氧诱导因子-1α;OXPHOS,线粒体氧化磷酸化;TNF-α,肿瘤坏死因子-α;TGF-β1,转化生长因子-β1;IL,白细胞介素。 图 1 肠道菌群经ROS/HIF-1α途径对巨噬细胞极化平衡的调控环节图 Fig. 1 Regulatory pathway diagram of intestinal flora on macrophage polarization balance via the ROS/HIF-1α pathway |
肠道菌群与中医学中的“脾”关系密切,可归属于“脾”的研究范畴,其结构组成可受到季节气候变化的影响而表现出一定的季节性特征,笔者团队进而提出肠道菌群可作为脾与长夏相通应的中介物质。长夏病理情况下,紊乱的肠道菌群可经过ROS/ HIF-1α途径调控机体的免疫反应发生异常改变,即巨噬细胞M1/M2的极化失衡,进而诱导UC的发生发展。
上述免疫调节功能随季节更迭而出现的节律性变动,为阐释长夏UC高发提供了潜在的免疫学依据。综合全文讨论,笔者团队提出“脾应长夏”的科学内涵可解释为1条与UC季节性发作密切相关的“肠道菌群—ROS/HIF-1α—巨噬细胞M1/M2极化”调控链。长夏温湿外令驱动菌群结构发生节律性重塑,差异菌群通过ROS/HIF-1α信号的时间性改变,调控巨噬细胞极化方向,从而奠定UC季节发作的免疫病理基础。这是基于“脾应长夏”探讨UC等消化系统疾病发病的生理病理基础之一。然而目前还亟待更多的基础研究与临床试验进行更深入的研究。
| [1] |
胡正芬, 刘永涛, 郑红斌. 从《内经》时脏的确立看脾主长夏的主导地位[J]. 辽宁中医药大学学报, 2007, 9(5): 56-57. |
| [2] |
中国中西医结合学会消化系统疾病专业委员会. 溃疡性结肠炎中西医结合诊疗共识意见(2017年)[J]. 中国中西医结合消化杂志, 2018, 26(2): 105-111, 120. |
| [3] |
BAI A P, GUO Y, SHEN Y H, et al. Seasonality in flares and months of births of patients with ulcerative colitis in a Chinese population[J]. Digestive Diseases and Sciences, 2009, 54(5): 1094-1098. DOI:10.1007/s10620-008-0453-1 |
| [4] |
GLAPA-NOWAK A, SZCZEPANIK M, KWIECIE? J, et al. Insolation and disease severity in paediatric inflammatory bowel disease-a multi-centre cross-sectional study[J]. Journal of Clinical Medicine, 2020, 9(12): 3957. DOI:10.3390/jcm9123957 |
| [5] |
王嘉源, 董卫国. 炎症性肠病患者入院季节性规律研究[J]. 胃肠病学和肝病学杂志, 2021, 30(9): 989-992. |
| [6] |
蒲晓田, 马淑然, 陈玉萍, 等. 关于中医"天人相应"理论内涵的探讨[J]. 中医杂志, 2012, 53(23): 1984-1986. |
| [7] |
覃骊兰, 马淑然, 刘燕池. 中医"脾主长夏"理论内涵的探讨[J]. 世界科学技术-中医药现代化, 2013, 15(8): 1815-1820. |
| [8] |
MURRAY P J. Macrophage polarization[J]. Annual Review of Physiology, 2017, 79: 541-566. DOI:10.1146/annurev-physiol-022516-034339 |
| [9] |
ZHOU F, MEI J T, HAN X G, et al. Kinsenoside attenuates osteoarthritis by repolarizing macrophages through inactivating NF-κB/MAPK signaling and protecting chondrocytes[J]. Acta Pharmaceutica Sinica B, 2019, 9(5): 973-985. DOI:10.1016/j.apsb.2019.01.015 |
| [10] |
MOMMERT S, JAHN M, SCHAPER-GERHARDT K, et al. Expression of histamine receptors H2R and H4R are predominantly regulated via the IL-4/IL-13 receptor type Ⅱ on human M2 macrophages[J]. Allergy, 2021, 76(9): 2886-2890. DOI:10.1111/all.14979 |
| [11] |
刘艺, 刘源, 孙志其, 等. 中药及有效成分靶向M1/M2巨噬细胞极化平衡治疗炎症性肠病的研究进展[J]. 中国实验方剂学杂志, 2024, 30(2): 276-286. |
| [12] |
PAN X H, ZHU Q, PAN L L, et al. Macrophage immunometabolism in inflammatory bowel diseases: From pathogenesis to therapy[J]. Pharmacology & Therapeutics, 2022, 238: 108176. |
| [13] |
FOGELSON K A, DORRESTEIN P C, ZARRINPAR A, et al. The gut microbial bile acid modulation and its relevance to digestive health and diseases[J]. Gastroenterology, 2023, 164(7): 1069-1085. DOI:10.1053/j.gastro.2023.02.022 |
| [14] |
ZHENG D P, LIWINSKI T, ELINAV E. Interaction between microbiota and immunity in health and disease[J]. Cell Research, 2020, 30(6): 492-506. DOI:10.1038/s41422-020-0332-7 |
| [15] |
孔凡浩, 闫慧, 李颖, 等. 肠黏膜屏障功能与消化系统疾病关系的研究进展[J]. 中国医药科学, 2023, 13(3): 59-62. |
| [16] |
郑昊龙, 陈丝, 宋囡, 等. 脾虚模型大鼠肠道菌群分布及时效性研究[J]. 中医杂志, 2020, 61(14): 1262-1267. |
| [17] |
曹静, 查安生. 四君子汤对溃疡性结肠炎大鼠肠道菌群调节作用研究[J]. 中医药临床杂志, 2019, 31(1): 102-104. |
| [18] |
BAI J, HU Y, BRUNER D W. Composition of gut microbiota and its association with body mass index and lifestyle factors in a cohort of 7-18 years old children from the American Gut Project[J]. Pediatric Obesity, 2019, 14(4): e12480. DOI:10.1111/ijpo.12480 |
| [19] |
黑利娟, 马士芬, 徐俊荣. 昼夜节律与胃肠道动力及肠道菌群的关系[J]. 国际消化病杂志, 2018, 38(4): 230-232. |
| [20] |
刘雷蕾, 都国文, 马淑然. 基于16SrDNA扩增子测序及靶向代谢组学探究"脾应长夏"的物质基础[J]. 中华中医药杂志, 2020, 35(5): 2529-2534. |
| [21] |
LIU J, SONG Y, LU X G, et al. Seasonal variation influences on intestinal microbiota in rats[J]. Current Microbiology, 2018, 75(8): 1006-1010. DOI:10.1007/s00284-018-1480-6 |
| [22] |
王菁. 基于"脾恶湿"探讨湿因素对大鼠各肠段免疫因子和肠道菌群的影响[D]. 北京: 北京中医药大学, 2014.
|
| [23] |
CAO Y N, FENG L J, LIU Y Y, et al. Effect of Lactobacillus rhamnosus GG supernatant on serotonin transporter expression in rats with post-infectious irritable bowel syndrome[J]. World Journal of Gastroenterology, 2018, 24(3): 338-350. DOI:10.3748/wjg.v24.i3.338 |
| [24] |
CHESTKOV I V, JESTKOVA E M, ERSHOVA E S, et al. ROS-induced DNA damage associates with abundance of mitochondrial DNA in white blood cells of the untreated schizophrenic patients[J]. Oxidative Medicine and Cellular Longevity, 2018, 2018: 8587475. DOI:10.1155/2018/8587475 |
| [25] |
靳鑫, 倪田根, 王宁, 等. PGC-1α通过线粒体介导巨噬细胞极化状态的机制研究[J]. 第三军医大学学报, 2019, 41(1): 56-62. |
| [26] |
LI R, ZHOU R, WANG H, et al. Gut microbiota-stimulated cathepsin K secretion mediates TLR4-dependent M2 macrophage polarization and promotes tumor metastasis in colorectal cancer[J]. Cell Death and Differentiation, 2019, 26(11): 2447-2463. DOI:10.1038/s41418-019-0312-y |
| [27] |
HOSANG L, CANALS R C, VAN DER FLIER F J, et al. The lung microbiome regulates brain autoimmunity[J]. Nature, 2022, 603(7899): 138-144. DOI:10.1038/s41586-022-04427-4 |
| [28] |
BRAVERMAN J, STANLEY S A. Nitric oxide modulates macrophage responses to Mycobacterium tuberculosis infection through activation of HIF-1α and repression of NF-κB[J]. Journal of Immunology, 2017, 199(5): 1805-1816. DOI:10.4049/jimmunol.1700515 |