 2026, Vol. 43
2026, Vol. 43文章信息
- 耿子瑶, 舒容丽, 李诗琦, 等.
- GENG Ziyao, SHU Rongli, LI Shiqi, et al.
- 脓毒症中性粒细胞多模态死亡方式的分子机制及中医药干预策略研究进展
- The regulatory mechanism of multimodal death of neutrophils in the pathological process of sepsis
- 天津中医药, 2026, 43(1): 119-128
- Tianjin Journal of Traditional Chinese Medicine, 2026, 43(1): 119-128
- http://dx.doi.org/10.11656/j.issn.1672-1519.2026.01.18
-
文章历史
- 收稿日期: 2025-08-05

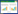



脓毒症是由宿主对感染的免疫反应失调所致的危及生命的器官功能障碍[1]。脓毒症的病死率和发病率都很高,全球疾病负担研究显示,2017年全球共有4 890万例新发脓毒症,死亡总人数共有1100万例,占当年全球死亡总人数的19.7%[2]。脓毒症可由各种损伤(外伤、烧伤、感染等)引起,在复杂的病理生理过程中,过度炎症反应和免疫调节失衡始终是其核心病理特征[3],免疫调节治疗成为研究的一个热点。
中性粒细胞是人体免疫系统中的重要组成部分,属于先天免疫反应的第一道防线。它们能够迅速迁移到感染或损伤部位,通过吞噬作用、释放抗菌物质以及形成中性粒细胞胞外陷阱(NETs)等多种方式来清除病原体[4]。脓毒症患者的中性粒细胞失调可引发严重后果:一方面,异常死亡的中性粒细胞会诱发免疫相关的器官损伤;另一方面,机体防御能力下降会增加院内感染风险。这种双重作用使得中性粒细胞从原本的“抗菌卫士”转变为加剧组织损伤和器官衰竭的“帮凶”。在脓毒症微环境中,中性粒细胞不仅表现出凋亡延迟,还可能因外部病理刺激而发生死亡方式转变[如坏死性凋亡、中性粒细胞胞外捕获物(NETosis)等][5],进而触发过度的炎症反应。更复杂的是,不同死亡途径之间存在交叉对话[6],某些关键分子甚至可能同时参与多种死亡程序的调控。
中医学对于脓毒症并没有明确的辨证理论,但从其临床症状和演变过程来看,脓毒症属于“温病” “热毒证”范畴,其病机特点为“正虚邪盛”“毒瘀互结”。在《黄帝内经》中就有“正气存内,邪不可干”的论述,强调机体免疫防御功能的重要性。中医药在调节免疫平衡、解毒化瘀方面具有独特优势。大量临床和实验研究表明,中药复方及活性成分能够通过多靶点、多途径调控中性粒细胞的死亡,从而减轻炎症反应、改善微循环障碍,这为脓毒症的治疗提供了新思路。
基于此,文章系统综述了中性粒细胞多模态死亡方式在脓毒症发生发展中的作用机制,重点探讨了凋亡抑制、NETosis异常激活、焦亡与炎症小体活化等关键病理环节。同时,阐述了中药复方(如血必净注射液、宣肺败毒方等)和单药成分(如甘草甜素、雷公藤甲素等)通过调节中性粒细胞死亡治疗脓毒症的研究进展,为深入理解脓毒症的免疫病理机制和开发新型治疗策略提供理论依据。

|
| 注:图片使用Procreate、Microsoft PowerPoint及Adobe Illustrator 2022绘制。TNFR1,肿瘤坏死因子受体1;FAS,肿瘤坏死因子受体超族,成员6;TRAIL,肿瘤坏死因子相关凋亡诱导配体;TRADD,肿瘤坏死因子受体相关死亡结构域蛋白;RIPK1,受体相互作用蛋白激酶1;Caspase-3,含半胱氨酸的天冬氨酸蛋白水解酶-3;Caspase-8,含半胱氨酸的天冬氨酸蛋白水解酶-8;Caspase-9,含半胱氨酸的天冬氨酸蛋白水解酶-9;BAX,B细胞淋巴瘤-2(BCL-2)相关X蛋白;BAK,BCL-2同源拮抗剂/杀伤因子;MCL-1,髓样细胞白血病序列1;NADPH,烟酰胺腺嘌呤二核苷酸磷酸(还原型辅酶Ⅱ);ROS,活性氧;DNA,脱氧核糖核酸;DAMPs,损伤相关分子模式;PAMPs,病原体相关分子模式;PRRs,模式识别受体;NLRP3,NOD样受体热蛋白结构域相关蛋白3;NLRC4,NOD样受体CARD结构域包含蛋白4;GSDMD,Gasdermin D蛋白;Caspase-1,含半胱氨酸的天冬氨酸蛋白水解酶-1;Caspase-11,含半胱氨酸的天冬氨酸蛋白水解酶-11;IL-1β,白细胞介素-1β;IL-18,白细胞介素-18;LPS,脂多糖。 图 1 中性粒细胞多模态死亡方式示意图 Fig. 1 Schematic diagram of multimodal death mode of neutrophils |
中性粒细胞作为免疫系统的重要组成部分,在生理状态下离开外周循环后会通过程序性凋亡途径发生有序的细胞死亡,这一机制可有效防止过度炎症反应的发生[7]。然而在脓毒症等病理状态下,过度激活的中性粒细胞可发生以细胞膜完整性丧失为特征的坏死性死亡(Necrosis)。该非程序性的细胞死亡方式导致细胞内蛋白水解酶(如弹性蛋白酶、组织蛋白酶等)、活性氧自由基及促炎因子等大量释放。这种失控的细胞内容物外泄不仅可直接造成周围组织损伤,更通过激活局部免疫细胞促进炎症级联放大,形成恶性循环[1]。坏死性中性粒细胞释放的DAMPs是介导其病理效应的关键介质,主要包括高迁移率族蛋白B1(HMGB1)、尿酸、热休克蛋白、DNA-染色质复合物和抗菌肽等[5]。这些DAMPs可被称为模式识别受体(PRR)的特定受体识别,从而启动免疫应答。其中Toll样受体(TLR)、RIG-I样受体(RLR)和NOD样受体(NLR)等可通过激活相关抗炎通路如NF-κB、丝裂原活化蛋白激酶(MAPK)等,诱导细胞因子和干扰素的产生,进而加剧炎症反应。值得注意的是,中性粒细胞坏死并非以直接杀菌为主要目的,而是作为一种机制,通过将颗粒内容物释放到细胞外环境中,促进免疫细胞的浸润,形成正反馈调节环路,导致炎症反应持续放大[8]。尽管中性粒细胞坏死在脓毒症中的确切分子机制尚未完全阐明,但现有研究表明,在抗微生物免疫应答过程中,中性粒细胞可发生坏死性死亡[1]。然而,目前关于脓毒症患者全身循环及组织中坏死性中性粒细胞的具体比例及其临床意义,仍需进一步研究证实。
2 凋亡细胞凋亡(Apoptosis)是由基因调控的主动、有序的细胞自我消亡过程。在生理状态下,正常成年人每天约产生1 011个成熟的中性粒细胞,而凋亡机制则通过有序清除衰老细胞来维持中性粒细胞数量的动态平衡[9-10]。这一过程对于炎症消退至关重要,因其能在不释放细胞毒性物质的前提下,有效清除炎症部位积聚的中性粒细胞,从而避免组织损伤[11]。然而,研究发现脓毒症可显著破坏这一生理过程。体外实验显示,静息培养的中性粒细胞24 h后约50% 呈现典型凋亡形态特征,而脓毒症患者来源的中性粒细胞凋亡率仅为5%~10%[1]。在急性肺损伤(ALI)动物模型中,无论是通过盲肠结扎穿刺(CLP)诱导的脓毒症模型,还是内毒素血症诱导的损伤模型,损伤后24 h内肺组织中性粒细胞凋亡率均呈现显著下降趋势[1, 12]。更值得关注的是,脓毒症患者血浆可显著延缓健康供者中性粒细胞的凋亡进程[13]。且临床研究证实,中性粒细胞凋亡抑制程度与脓毒症患者的疾病严重程度呈负相关,提示其可能作为评估病情进展的重要生物学指标[14]。
中性粒细胞凋亡延迟的主要原因可能与抗凋亡因子的激活有关[15]。临床和实验研究均表明,重症脓毒症患者外周血中性粒细胞中抗凋亡蛋白髓细胞白血病1(Mcl-1)的表达水平显著升高[16]。深入研究发现,多种信号通路参与调控脓毒症状态下中性粒细胞的凋亡延迟:循环中的促炎介质,如脂多糖(LPS)和补体成分5a(C5a),可通过激活中性粒细胞外调节蛋白激酶(ERK)1/2和磷酸肌醇-3激酶(PI3K)信号通路,进而上调抗凋亡蛋白(Bcl-xl)的表达,同时抑制促凋亡蛋白(Bim)的表达[17];脓毒症患者中性粒细胞中NF-κB的异常激活与半胱天冬氨酸蛋白酶(Caspase)-3、Caspase-9活性降低以及线粒体跨膜电位维持密切相关,这些改变共同导致细胞凋亡延迟[1, 18];MNDA作为中性粒细胞凋亡的重要调控因子,其核质转位过程对Mcl-1的降解具有关键作用,在脓毒症状态下,Caspase-8活性降低导致MNDA切割/重新定位障碍,伴随Mcl-1的异常积累,最终抑制中性粒细胞凋亡[19]。最新研究发现,腺苷A2A受体(A2AR)可通过抑制自噬过程,阻断Caspase-8、Caspase-3和聚腺苷二磷酸核糖聚合酶(PARP)信号通路,从而维持中性粒细胞的存活[20]。在治疗策略方面,研究人员近期开发了一种基于阿霉素(DOX)偶联蛋白纳米颗粒(NPs)的靶向递送系统。该系统能够特异性靶向炎症部位的中性粒细胞,通过诱导细胞凋亡显著提高脓毒症小鼠的存活率[1],这一突破为脓毒症治疗提供了新的思路。尽管目前对脓毒症中性粒细胞凋亡抑制机制已有一定认识,但其精确的调控网络仍需进一步阐明。虽然其凋亡受到抑制,但是还会经历一些其他类型的细胞死亡,外部环境的病理刺激或变化会诱导中性粒细胞死亡方式的转变,并导致压倒性的炎症反应[5-6]。
3 NETosisNETosis自2004年被阐明以来,成为先天免疫领域的研究热点。作为一种独特的程序性死亡方式,NETosis通过释放包有瓜氨酸化组蛋白、弹性蛋白酶、髓过氧化物酶(MPO)和组织蛋白酶G等的DNA网状结构,发挥其限制细菌传播和消灭病原体的作用。然而在脓毒症病理进程中,NETs可被看作“双刃剑”:在感染的早期阶段,适度的NETs形成能够有效限制病原体扩散并促进其清除;而重症阶段异常增多的NETs则会成为诱导血管内血栓形成、弥散性血管内凝血(DIC)和多器官功能障碍(MODS)的关键因素。
脓毒症引发的内皮功能障碍与NETs过度释放密切相关。过度的中性粒细胞活化和NETs释放可通过病原体识别受体(如TLR)诱导内皮细胞转向促炎和促凝血表型[1]。研究表明,作为NETs的重要组分,中性粒细胞弹性蛋白酶(NE)可通过蛋白水解作用调控多种血管生成关键生长因子(如血管内皮生长因子VEGF和血小板衍生生长因子PDGF)的活性[21]。1项研究发现阻断一种生长因子可以补偿性地上调其他生长因子的表达并再次刺激血管生成[22]。当VEGF被阻断时,其他生长因子(如成纤维细胞生长因子FGF)的表达可能会升高,从而通过FGF-MYC-HK2轴再次促进糖酵解,破坏内皮细胞代谢平衡[23]。此外,NETs可通过破坏糖萼、降解紧密连接蛋白以及直接毒性作用促进内皮功能障碍。首先,中性粒细胞和NETs可以降解内皮细胞表面的糖萼(由糖胺聚糖、蛋白聚糖及糖蛋白构成的保护性基质网)并增加内皮通透性[1],内皮屏障塌陷,导致脓毒症晚期微循环血流受损、组织灌注不足和危及生命的器官衰竭[1]。MPO、基质金属蛋白酶(MMP)和其他含NETs蛋白酶的酶消化作用会导致糖萼降解[24-25]。临床已在脓毒症患者血浆中检测到糖萼脱落标志物(如Syndecan-1、透明质酸)[26],且其水平与病情严重程度呈正相关[27]。尽管MMP-9的生物学功能尚存争议,但临床研究数据显示,MMP-9的活性形式仅存在于重症脓毒症患者中[28]。其次,许多由中性粒细胞和NETs诱导的炎性细胞因子,如TNF-α、白细胞介素(IL)-6和IL-8,可直接损害糖萼[29]。脓毒症期间内皮旁通透性增加主要是由于连接处裂解。TNF-α可以通过激活NF-κB通路来破坏内皮细胞胞间连接处的Claudin-5[30]。最后,NETs中的组蛋白和MPO可直接介导内皮细胞毒性,以剂量依赖方式诱导细胞死亡[31]。在LPS诱导的脓毒症小鼠模型中,PAD-NET信号通路激活导致的NETs大量生成,已被证实可显著增加肺血管内皮通透性[32]。
凝血系统的激活是脓毒症重要的免疫防御反应,可以限制微生物的传播[33]。然而,NETs的过度产生会打破凝血级联反应的动态平衡。研究表明,血小板活化后可诱导中性粒细胞释放NETs,进而触发血栓形成[1]。NETs的持续释放不仅会在血液中引发血栓,加剧局部组织损伤,还与弥漫性血管内凝血病的病理进程密切相关[1]。炎症部位携带组织因子的NETs可导致凝血级联反应的局部激活[34]。NETs的各种蛋白水解成分可通过多种途径激活血管内凝血。其中,丝氨酸蛋白酶(如NE和组织蛋白酶G)可通过降解抗凝血酶等生理凝血抑制剂加速凝血进程;NETs的核心成分染色质和组蛋白,更是强效的凝血启动剂[2]。此外,NETs独特的网状结构可为血栓形成提供物理支架[1],最终导致微血管系统阻塞,引发组织缺血和损伤,甚至进展为多器官功能衰竭,危及生命。靶向NETs的治疗策略可能成为脓毒症治疗的新突破口。此外,NETosis的发生并非孤立事件,其与自噬、焦亡等过程存在显著交叉。
4 自噬自噬(Autophagy)是一种稳态机制,细胞通过自噬分解自身损伤或老化的细胞器、清除入侵的病原体和聚集的错误折叠蛋白,实现代谢废物循环利用与营养物质再分配[35]。这一过程始于双层膜结构自噬体的形成,随后自噬体与溶酶体融合形成自噬溶酶体,其内容物被溶酶体内的酶分解,分解产物随后被释放回细胞内环境,供细胞再利用,如能量产生或新分子的合成[36]。自噬是细胞在应激条件下的一种适应性保护机制,而非单纯的细胞死亡途径。目前,许多关于中性粒细胞自噬与脓毒症的研究表明,自噬具有缓解脓毒症的作用。例如,自噬相关基因(ATG)蛋白是自噬的关键调节因子,是雷帕霉素复合物1(mTORC1)机制靶标的下游元件。ATG7缺陷导致IL-1β产生增加和细胞焦亡的发生,导致脓毒症假单胞菌小鼠模型中病原体清除受损、存活率降低和肺损伤加重[37]。一些研究表明,激活的自噬可以改善由脓毒症诱发的肺损伤[38-40]、肾损伤[41]和心脏损伤[42]。
NETs的形成受多种因素调控,其中自噬在这一过程中的具体作用机制尚存在争议[43-44]。为了更好地解释NETs在不同疾病中的可变蛋白载量和作用,研究者提出了“two-hit”模型。该模型认为,炎症微环境中的多种刺激因子形成的初次刺激(first hit)可促使中性粒细胞上调疾病相关蛋白的表达;而2次刺激(second hit)则触发NETs的形成,通过其特有的支架结构实现胞内蛋白的定向释放,这一过程在增强宿主抗菌能力的同时,也可能加剧炎症反应[1, 45]。多项证据表明自噬可能充当关键的“second/NETotic-hit”,负责调控NETs相关效应分子的胞外递送过程[46, 1]。来自脓毒症患者的临床数据显示,其中性粒细胞不仅呈现自噬活性增强的特征,还对后续佛波醇12-肉豆蔻酸酯13-乙酸(PMA)刺激诱导的NETosis表现出异常敏感性,这种双向调控关系在小鼠模型中进一步得到验证——适度的自噬诱导可通过NETs依赖性机制显著提高脓毒症动物的生存率[47]。然而,自噬驱动的NETs的过量产生也会导致组织损伤,从而加速疾病进展。一项临床研究表明,NETs形成增加与脓毒症诱导的弥散性血管内凝血(DIC)发病率和病死率呈显著正相关[48]。在分子机制层面,研究发现自噬与NETs形成共同受PI3K-AKT-mTOR信号轴调控,利用HL-60细胞分化体系的研究表明,当敲除该通路关键负调控因子PTEN(兼具自噬诱导功能)时,中性粒细胞样细胞经PMA刺激后产生的NETs显著减少[49]。
然而,在Atg5基因敲除小鼠模型中,虽然中性粒细胞的自噬活性被显著抑制,但其NETs释放能力却未受明显影响[50]。这一发现直接挑战了自噬在NETosis过程中不可或缺的传统观点。更为复杂的是,有研究团队通过系统的实验证实,NETs的形成过程同时涉及NADPH氧化酶依赖性ROS的产生和自噬激活这两个关键事件,但这两个通路在功能上表现出明显的独立性,可以平行而非序贯地促进NETosis的发生[51]。
5 焦亡细胞焦亡(Pyroptosis)是一种程序性细胞死亡方式,具有高度促炎性,是依赖于Gasdermin家族蛋白形成质膜(PM)膜孔的可调控过程,Gasdermin-D(GSDMD)蛋白经Caspase切割后,释放具有膜打孔活性的N端片段(N-GSDMD)。N-GSDMD可识别并结合细胞膜上的磷脂类分子并打孔,促使细胞膜裂解,细胞内容物释放,最终导致细胞死亡[1, 52]。该过程可通过经典和非经典途径激活:经典途径由病原相关分子模式(PAMPs)/损伤相关的分子模式(DAMPs)触发,通过炎症小体(如NLRP3)激活Caspase-1,进而切割GSDMD并促进IL-1β/IL-18成熟[53];非经典途径则由胞内LPS直接激活Caspase-4/5/11切割GSDMD[54]。在中性粒细胞中,细胞焦亡促进先天免疫主要有两种途径:1)消除病原菌的细胞内复制生态位;2)通过非经典分泌途径释放IL-1家族细胞因子,放大宿主对微生物感染或组织损伤的有益炎症反应。
在正常的生理过程中,细胞焦亡在宿主防御病原感染中起着关键作用。然而,过度的细胞焦亡会导致压倒性和持续的炎症反应,这与脓毒症的发病机制有关。在脓毒症模型中,中性粒细胞中N-乙酰转移酶10(NAT10)的表达水平显著降低。通过基因工程技术特异性上调中性粒细胞NAT10表达,可有效调控ULK1-STING-NLRP3信号轴,从而抑制中性粒细胞过度焦亡,显著改善脓毒症小鼠的生存率[55]。此外,在脓毒症发病过程中,中性粒细胞焦亡的关键效应蛋白(包括Caspase-1/11和GSDMD)发挥着重要的调控作用。有研究发现,内源性Caspase-1的激活会显著降低多种微生物所致脓毒症模型的存活率,并促进炎性细胞因子的过度释放。值得注意的是,通过基因敲除Caspase-1/11可有效改善中性粒细胞的吞噬功能[56]。另有研究进一步证实,Caspase-11是中性粒细胞发生焦亡的必要条件,同时也是清除伯克霍尔德菌感染的关键因素[57]。
此外,在脓毒症微环境中,GSDMD介导的中性粒细胞焦亡与NETosis密切相关。Caspase - 11与GSDMD通过协同调控核膜透化及组蛋白降解过程,进而介导NETs的形成与释放。在CLP诱导的脓毒症模型中,Caspase-11敲除(而非Caspase-1敲除)能有效抑制NETs释放,从而缓解脓毒症症状[58]。Caspase-11激活促进暴露于尿酸钠的中性粒细胞以及来自脓毒症小鼠和患者的中性粒细胞的NETosis[1, 59],表明GSDMD诱导的细胞质膜破裂可能导致NETs释放。中性粒细胞中GSDMD裂解介导的NETs释放机制也可能是急性呼吸窘迫综合征(ARDS)发病的关键环节[60]。这种由GSDMD介导的细胞死亡方式在先天免疫中具有重要意义,它不仅能够直接清除被病原体感染的细胞,还能通过释放细胞内容物来警示免疫系统。
6 中医药调控中性粒细胞死亡的潜力 6.1 中药复方及其制剂血必净注射液是一种基于传统方剂血府逐瘀汤改良而成的复方静脉注射剂,由天津市第一中心医院王金达教授团队研发。该制剂由5种中药成分组成:芍药(Paeonia lactiflora Pall.)、川芎(Ligusticum chuanxiong Hort.)、红花(Carthamus tinctorius L.)、当归[Angelica sinensis(Oliv.)Diels]和丹参(Salvia miltiorrhiza Bunge)。基于细菌-毒素-炎症三联治疗理论,血必净注射液在临床上主要用于脓毒症和脓毒性休克的治疗,其作用机制包括抑制血小板过度活化和病理性血栓形成[61]。研究发现,NETs的过度释放是导致脓毒症患者器官功能衰竭和死亡的关键因素。在临床相关的多微生物感染模型中,血必净注射液能够显著降低瓜氨酸化组蛋白H3(CitH3)、MPO和NE的表达水平。机制研究表明,焦亡相关蛋白GSDMD介导NETs的形成[62],血必净注射液通过抑制GSDMD相关基因的表达,阻断GSDMD介导的NETs形成途径[63]。进一步分析发现,血必净注射液中的3种主要活性成分——羟基红花黄色素A、红花黄色素A和无水红花黄色素B,能有效降低血浆中MPO、组蛋白及MPO-DNA复合物的水平[64-65]。
宣肺败毒方是中国在抗击新型冠状病毒感染(COVID-19)疫情期间研发的中药复方制剂,由张伯礼院士团队在传统方剂基础上优化而成,被纳入《新型冠状病毒肺炎诊疗方案》。研究发现,宣肺败毒汤可通过调控CXC趋化因子配体2(CXCL2)/ CXC趋化因子受体2(CXCR2)信号通路,影响NETosis[66]。
四妙勇安汤具有清热解毒、活血止痛的功效,主要用于治疗热毒炽盛导致的脱疽,在临床上可用于治疗脓毒症[67]。体外实验表明,20% 浓度的四妙勇安汤含药血清能够显著抑制中性粒细胞迁移,同时促进中性粒细胞细胞凋亡,并有效降低ROS生成和NETs的形成[68]。
热毒宁注射液具有疏风解表,清热解毒之功效。其在临床上常用于治疗上呼吸道感染,并且对多重耐药菌引发的呼吸道感染患者有较好的临床效果。研究发现,热毒宁注射液通过调节MAPK途径抑制NETosis来改善LPS诱导的ALI[69]。
紫雪散作为清热解毒、开窍止痉经典方剂,临床广泛用于急症热病及危重症治疗。最新研究发现,紫雪散通过抑制STING介导的血小板可溶性N-乙基马来酰亚胺敏感因子附着蛋白受体(SNAREs)复合物的组装来减弱P-选择素的分泌,最终抑制NETs的形成[70]。
荆银固表方具有疏风解表、清热解毒、芳香避秽、培土生金的功效,已广泛用于治疗急性呼吸道感染。该方可通过抑制中性粒细胞中的ERK/ROS轴来抑制NETs形成、缓解炎症反应,从而减轻了流感后金黄色葡萄球菌感染小鼠的ALI[71]。
6.2 单药及其成分姜黄二酮(Curdione)是中药姜黄(Curcuma longa)的主要活性成分之一,具有广泛的药理活性,包括抗炎、抗氧化及抑制血小板聚集等作用。研究表明,Curdione可通过抑制血小板介导的中性粒细胞募集与浸润,减少NETs的形成,并发挥其抗炎及抗氧化特性,从而有效减轻脓毒症诱导的肺损伤[72]。
甘草甜素(GL)是从光果甘草(Glycyrrhiza glabra L.)中提取的一种天然三萜类化合物,具有抗炎、抗病毒和免疫调节等多种药理活性。最新研究表明,GL通过多靶点作用机制对脓毒症及其并发症发挥显著保护作用:首先,HMGB1通过调控中性粒细胞的活化、染色质解聚及炎症信号放大,成为NETs形成的核心调控分子,GL能特异性结合HMGB1,阻断其介导的炎症级联反应,显著降低IL-6和TNF-α等促炎因子的释放[73-74];其次,在CLP模型中,GL通过下调HMGB1/TLR9/MyD88信号通路抑制NETs形成,从而减轻肺损伤并提高生存率[75]。
洋川芎内酯I(SEI)是从中药川芎(Ligusticum chuanxiong Hort.)中分离得到的一种特征性邻苯二甲酸内酯类化合物,具有显著的镇痛、抗血栓形成和抗炎等多重药理活性[76]。作为血必净注射液的关键组分之一被广泛应用于脓毒症的临床治疗。研究表明,SEI能够有效降低TNF-α、IL-1β和IL-6等炎性细胞因子的含量,并通过调控CLP诱导的脓毒症小鼠的血浆和肺组织中c-Jun氨基末端激酶(JNK)和细胞外信号调节激酶(ERK)信号通路的活化来抑制MPO-DNA复合物的水平[77]。
连翘苷B(Forsythiaside B)是从中药连翘(Forsythia suspensa)中提取的一种苯乙醇苷类化合物,具有抗炎、心脏保护和神经保护等多种药理活性。最新研究表明,连翘苷B可通过特异性抑制外周血中性粒细胞肽酰基精氨酸脱亚胺酶4(PAD4)依赖性NETs的形成,显著改善CLP诱导的脓毒症大鼠模型的凝血功能障碍[78]。
粉防己碱(Tetrandrine)是一种从中药粉防己(Stephania tetrandra S. Moore)中提取的双苄基异喹啉类生物碱。在体外,tetrandrine抑制PMA诱导的NETosis [79],同时降低中性粒细胞IL-6的释放。
穿心莲内酯(Andrographolide)是从中药穿心莲(Andrographis paniculata)中分离得到的一种拉布丹型二萜类活性成分,其磺化衍生物穿心莲内酯磺酸盐(Andrographolide sulfonate)通过抑制中性粒细胞衍生的蛋白酶(如MPO、NE和组织蛋白酶G)的表达来减轻LPS诱导的肺损伤[80]。
雷公藤甲素(Triptolide)是从中药雷公藤(Tripterygium wilfordii)中分离得到的环氧二萜内酯类化合物,可抑制关节炎小鼠中性粒细胞的迁移、自噬和内分泌性[81]。该化合物对健康志愿者外周血中性粒细胞具有显著的抗NETosis作用,且这种作用主要与其抗炎特性相关,而非通过调节氧化应激途径实现[81-82]。雷公藤甲素的抗炎活性还改善了LPS诱导的急性呼吸窘迫综合征小鼠肺泡高凝状态[83]。
白及多糖是从中药白及(Bletilla striata)假鳞茎中提取的一种葡甘聚糖类成分。研究表明,白及多糖可能通过调控PAD4通路维持中性粒细胞-巨噬细胞稳态,进而抑制NETs的形成,减轻NETs诱导的肺泡巨噬细胞焦亡,从而改善ARDS的病理进程[84]。
三七(Panax notoginseng)在抗炎与免疫调节治疗领域具有悠久的临床应用历史。其主要活性成分皂苷[包括Rh1、Rk3、Rh4、20(S)-Rg3和20(R)-Rg3]的含量随蒸制时间的延长而增加。研究发现,这些皂苷能抑制中性粒细胞活性,其中Rh1和20(R)-Rg3可抑制中性粒细胞迁移,而Rk3、Rh4和20(S)-Rg3不仅能抑制迁移,还能促进中性粒细胞凋亡[85]。
研究显示,SARS-CoV-2感染可诱发NETs形成、凝血功能异常及炎症性细胞因子风暴。聚乙二醇化纳米白蛋白结合(PNAB)甾体人参皂苷制剂(PNAB-人参皂苷Rg6和PNAB-人参皂苷Rgx365,能显著降低组蛋白H4及NETs相关标志物(如MPO和NE)的血清浓度[86]。
木犀草素(Luteolin)是一种广泛存在于多种药用植物中的天然黄酮类化合物,具有显著的抗衰老、抗炎及抗肿瘤等多重药理活性。研究证明木犀草素通过调节RAF1-MEK-1-ERK通路有效抑制人中性粒细胞炎症反应、弹性蛋白酶释放和NETosis[87]。在脓毒症小鼠中,木犀草素可降低血清中HMGB1的含量并减轻肝损伤[88]。
7 小结中性粒细胞作为免疫系统的重要组成部分,在脓毒症中扮演着至关重要的角色。尽管凋亡一直是研究的热点,但坏死、自噬、焦亡和NETosis在这一危急情况下同样发挥着重要作用[5]。在脓毒症致病过程中,中性粒细胞以上的“牺牲”行为是一把双刃剑:保护性机制方面,凋亡的有序发生可清除衰老细胞以限制过度炎症,NETosis释放的抗菌网状结构及焦亡介导的胞内病原体清除,构成抵御微生物侵袭的关键防线;损伤性机制方面,坏死导致的DAMPs暴发性释放、凋亡延迟引发的“衰老中性粒细胞”蓄积,以及NETosis单独或者焦亡/自噬-NETosis协同驱动的炎症-凝血级联放大,共同加剧血管内皮损伤、微循环障碍及多器官功能衰竭。这些死亡方式可能并非单独存在,因为这对于调节宿主免疫反应至关重要。中性粒细胞的死亡方式可能受病原体毒力因子、刺激的持续时间及强度、组织损伤程度及宿主免疫应答共同调控。此外,不同死亡方式之间的交互对话也可能构成脓毒症免疫紊乱的重要分子基础。例如,焦亡和自噬可能通过调节炎症小体活性和细胞内容物释放来影响NETosis的发生;而NETosis过程中释放的组蛋白和蛋白酶又可能反过来调控其他细胞死亡途径。
尽管脓毒症中性粒细胞死亡的机制仍有待阐明,针对中性粒细胞死亡的药物干预甚至很少,但目前中医药在调控中性粒细胞死亡方面已经展现出独特潜力。例如,复方制剂(如血必净注射液、宣肺败毒方等)和单药成分(如姜黄二酮、甘草甜素等)通过靶向关键信号通路(如GSDMD、HMGB1/ TLR9等),有效抑制NETosis和焦亡,减轻炎症反应和器官损伤。此外,目前中医药对中性粒细胞死亡的调控存在明显选择性,NETosis和凋亡是主要靶点,其他死亡方式需更多机制研究。
中医药调控中性粒细胞死亡是一种有希望的治疗方法。因此,未来的研究方向应该着重于:1)阐明不同细胞死亡方式在脓毒症不同阶段的具体贡献;2)开发能够精确调控特定细胞死亡途径的靶向治疗策略以扩大其保护作用;)探索中医药多3组分协同调控的分子机制。这些研究将为脓毒症的精准治疗提供新的思路和理论依据。
| [1] |
SINGER M, DEUTSCHMAN C S, SEYMOUR C W, et al. The third international consensus definitions for sepsis and septic shock (sepsis-3)[J]. JAMA, 2016, 315(8): 801-810. DOI:10.1001/jama.2016.0287 |
| [2] |
RUDD K E, JOHNSON S C, AGESA K M, et al. Global, regional, and national sepsis incidence and mortality, 1990-2017:Analysis for the Global Burden of Disease Study[J]. The Lancet, 2020, 395(10219): 200-211. DOI:10.1016/S0140-6736(19)32989-7 |
| [3] |
VAN DER POLL T, VAN DE VEERDONK F L, SCICLUNA B P, et al. The immunopathology of sepsis and potential therapeutic targets[J]. Nature Reviews Immunology, 2017, 17(7): 407-420. DOI:10.1038/nri.2017.36 |
| [4] |
LIEW P X, KUBES P. The neutrophil's role during health and disease[J]. Physiological Reviews, 2019, 99(2): 1223-1248. DOI:10.1152/physrev.00012.2018 |
| [5] |
IBA T, HASHIGUCHI N, NAGAOKA I, et al. Neutrophil cell death in response to infection and its relation to coagulation[J]. Journal of Intensive Care, 2013, 1(1): 13. DOI:10.1186/2052-0492-1-13 |
| [6] |
PéREZ-FIGUEROA E, áLVAREZ-CARRASCO P, ORTEGA E, et al. Neutrophils: Many ways to die[J]. Frontiers in Immunology, 2021, 12: 631821. DOI:10.3389/fimmu.2021.631821 |
| [7] |
HASLETT C. Granulocyte apoptosis and its role in the resolution and control of lung inflammation[J]. American Journal of Respiratory and Critical Care Medicine, 1999, 160(5 Pt 2): S5-11. |
| [8] |
NAYLOR E J, BAKSTAD D, BIFFEN M, et al. Haemophilus influenzae induces neutrophil necrosis: A role in chronic obstructive pulmonary disease?[J]. American Journal of Respiratory Cell and Molecular Biology, 2007, 37(2): 135-143. DOI:10.1165/rcmb.2006-0375OC |
| [9] |
MIWA K, ASANO M, HORAI R, et al. Caspase 1-independent IL1beta release and inflammation induced by the apoptosis inducer Fas ligand[J]. Nature Medicine, 1998, 4(11): 1287-1292. DOI:10.1038/3276 |
| [10] |
SANSONETTI P J, PHALIPON A, ARONDEL J, et al. Caspase-1 activation of IL-1beta and IL-18 are essential for Shigella flexneriinduced inflammation[J]. Immunity, 2000, 12(5): 581-590. DOI:10.1016/S1074-7613(00)80209-5 |
| [11] |
SAVILL J, FADOK V. Corpse clearance defines the meaning of cell death[J]. Nature, 2000, 407: 784-788. DOI:10.1038/35037722 |
| [12] |
WANG J F, WANG Y P, XIE J, et al. Upregulated PD-L1 delays human neutrophil apoptosis and promotes lung injury in an experimental mouse model of sepsis[J]. Blood, 2021, 138(9): 806-810. DOI:10.1182/blood.2020009417 |
| [13] |
JIMENEZ M F, WATSON R W, PARODO J, et al. Dysregulated expression of neutrophil apoptosis in the systemic inflammatory response syndrome[J]. Archives of Surgery, 1997, 132(12): 1263-1269. DOI:10.1001/archsurg.1997.01430360009002 |
| [14] |
FIALKOW L, FOCHESATTO FILHO L, BOZZETTI M C, et al. Neutrophil apoptosis: A marker of disease severity in sepsis and sepsis-induced acute respiratory distress syndrome[J]. Critical Care, 2006, 10(6): R155. DOI:10.1186/cc5090 |
| [15] |
WESCHE D E, LOMAS-NEIRA J L, PERL M, et al. Leukocyte apoptosis and its significance in sepsis and shock[J]. Journal of Leukocyte Biology, 2005, 78(2): 325-337. DOI:10.1189/jlb.0105017 |
| [16] |
LUAN Y Y, YAO Y M, XIAO X Z, et al. Insights into the apoptotic death of immune cells in sepsis[J]. Journal of Interferon & Cytokine Research, 2015, 35(1): 17-22. |
| [17] |
GUO R F, SUN L, GAO H W, et al. In vivo regulation of neutrophil apoptosis by C5a during sepsis[J]. Journal of Leukocyte Biology, 2006, 80(6): 1575-1583. DOI:10.1189/jlb.0106065 |
| [18] |
POWER C P, WANG J H, MANNING B, et al. Bacterial lipoprotein delays apoptosis in human neutrophils through inhibition of Caspase-3 activity: Regulatory roles for CD14 and TLR-2[J]. Journal of Immunology, 2004, 173(8): 5229-5237. DOI:10.4049/jimmunol.173.8.5229 |
| [19] |
MILOT E, FOTOUHI-ARDAKANI N, FILEP J G. Myeloid nuclear differentiation antigen, neutrophil apoptosis and sepsis[J]. Frontiers in Immunology, 2012, 3: 397. |
| [20] |
LIU Y W, YANG T, ZHAO L, et al. Activation of Adenosine 2A receptor inhibits neutrophil apoptosis in an autophagy-dependent manner in mice with systemic inflammatory response syndrome[J]. Scientific Reports, 2016, 6: 33614. DOI:10.1038/srep33614 |
| [21] |
EUSTACHE J H, TOHME S, MILETTE S, et al. Casting A wide net on surgery: The central role of neutrophil extracellular traps[J]. Annals of Surgery, 2020, 272(2): 277-283. DOI:10.1097/SLA.0000000000003586 |
| [22] |
EBOS J M L, KERBEL R S. Antiangiogenic therapy: Impact on invasion, disease progression, and metastasis[J]. Nature Reviews Clinical Oncology, 2011, 8(4): 210-221. DOI:10.1038/nrclinonc.2011.21 |
| [23] |
YU P C, WILHELM K, DUBRAC A, et al. FGF-dependent metabolic control of vascular development[J]. Nature, 2017, 545(7653): 224-228. DOI:10.1038/nature22322 |
| [24] |
ALI M M, MAHMOUD A M, LE MASTER E, et al. Role of matrix metalloproteinases and histone deacetylase in oxidative stress-induced degradation of the endothelial glycocalyx[J]. American Journal of Physiology Heart and Circulatory Physiology, 2019, 316(3): H647-H663. DOI:10.1152/ajpheart.00090.2018 |
| [25] |
MANCHANDA K, KOLAROVA H, KERKENPA? C, et al. MPO (myeloperoxidase) reduces endothelial glycocalyx thickness dependent on its cationic charge[J]. Arteriosclerosis, Thrombosis, and Vascular Biology, 2018, 38(8): 1859-1867. DOI:10.1161/ATVBAHA.118.311143 |
| [26] |
OSTROWSKI S R, HAASE N, MüLLER R B, et al. Association between biomarkers of endothelial injury and hypocoagulability in patients with severe sepsis: A prospective study[J]. Critical Care, 2015, 19(1): 191. DOI:10.1186/s13054-015-0918-5 |
| [27] |
SMART L, MACDONALD S P J, BURROWS S, et al. Endothelial glycocalyx biomarkers increase in patients with infection during Emergency Department treatment[J]. Journal of Critical Care, 2017, 42: 304-309. DOI:10.1016/j.jcrc.2017.07.001 |
| [28] |
G?DDN?S F P, SUTINEN M M, KOSKELA M, et al. Matrix-metalloproteinase-2, -8 and -9 in serum and skin blister fluid in patients with severe sepsis[J]. Critical Care, 2010, 14(2): R49. DOI:10.1186/cc8938 |
| [29] |
SCHMIDT E P, YANG Y M, JANSSEN W J, et al. The pulmonary endothelial glycocalyx regulates neutrophil adhesion and lung injury during experimental sepsis[J]. Nature Medicine, 2012, 18(8): 1217-1223. DOI:10.1038/nm.2843 |
| [30] |
CLARK P R, KIM R K, POBER J S, et al. Tumor necrosis factor disrupts claudin-5 endothelial tight junction barriers in two distinct NF-κB-dependent phases[J]. PLoS One, 2015, 10(3): e0120075. DOI:10.1371/journal.pone.0120075 |
| [31] |
SAFFARZADEH M, JUENEMANN C, QUEISSER M A, et al. Neutrophil extracellular traps directly induce epithelial and endothelial cell death: A predominant role of histones[J]. PLoS One, 2012, 7(2): e32366. DOI:10.1371/journal.pone.0032366 |
| [32] |
LIANG Y J, PAN B H, ALAM H B, et al. Inhibition of peptidylarginine deiminase alleviates LPS-induced pulmonary dysfunction and improves survival in a mouse model of lethal endotoxemia[J]. European Journal of Pharmacology, 2018, 833: 432-440. DOI:10.1016/j.ejphar.2018.07.005 |
| [33] |
ESMON C T, XU J, LUPU F. Innate immunity and coagulation[J]. Journal of Thrombosis and Haemostasis, 2011, 9(Suppl 1): 182-188. |
| [34] |
KAMBAS K, MITROULIS I, APOSTOLIDOU E, et al. Autophagy mediates the delivery of thrombogenic tissue factor to neutrophil extracellular traps in human sepsis[J]. PLoS One, 2012, 7(9): e45427. DOI:10.1371/journal.pone.0045427 |
| [35] |
GLICK D, BARTH S, MACLEOD K F. Autophagy: Cellular and molecular mechanisms[J]. The Journal of Pathology, 2010, 221(1): 3-12. DOI:10.1002/path.2697 |
| [36] |
BHATTACHARYA A, WEI Q, SHIN J N, et al. Autophagy is required for neutrophil-mediated inflammation[J]. Cell Reports, 2015, 12(11): 1731-1739. DOI:10.1016/j.celrep.2015.08.019 |
| [37] |
PU Q Q, GAN C P, LI R P, et al. Atg7 deficiency intensifies inflammasome activation and pyroptosis in Pseudomonas sepsis[J]. Journal of Immunology, 2017, 198(8): 3205-3213. DOI:10.4049/jimmunol.1601196 |
| [38] |
ZHAO H Y, CHEN H G, MENG X Y, et al. Autophagy activation improves lung injury and inflammation in sepsis[J]. Inflammation, 2019, 42(2): 426-439. DOI:10.1007/s10753-018-00952-5 |
| [39] |
AGUIRRE A, LóPEZ-ALONSO I, GONZáLEZ-LóPEZ A, et al. Defective autophagy impairs ATF3 activity and worsens lung injury during endotoxemia[J]. Journal of Molecular Medicine, 2014, 92(6): 665-676. DOI:10.1007/s00109-014-1132-7 |
| [40] |
ZUO Y L, DANG R, PENG H Y, et al. LL-37 exacerbates local inflammation in sepsis-induced acute lung injury by preventing mitochondrial DNA(mtDNA) degradation-induced autophagy[J]. Medical Science Monitor, 2019, 25: 6193-9203. DOI:10.12659/MSM.915298 |
| [41] |
ZHAO Y, FENG X J, LI B, et al. Dexmedetomidine protects against lipopolysaccharide-induced acute kidney injury by enhancing autophagy through inhibition of the PI3K /AKT /mTOR pathway[J]. Frontiers in Pharmacology, 2020, 11: 128. DOI:10.3389/fphar.2020.00128 |
| [42] |
HSIEH C H, PAI P Y, HSUEH H W, et al. Complete induction of autophagy is essential for cardioprotection in sepsis[J]. Annals of Surgery, 2011, 253(6): 1190-1200. DOI:10.1097/SLA.0b013e318214b67e |
| [43] |
YOUSEFI S, STOJKOV D, GERMIC N, et al. Untangling"netosis" from NETs[J]. European Journal of Immunology, 2019, 49(2): 221-227. DOI:10.1002/eji.201747053 |
| [44] |
YOUSEFI S, SIMON H U. NETosis-does it really represent nature's "suicide bomber"?[J]. Frontiers in Immunology, 2016, 7: 328. |
| [45] |
MITSIOS A, ARAMPATZIOGLOU A, ARELAKI S, et al. NETopathies? Unraveling the dark side of old diseases through neutrophils[J]. Frontiers in Immunology, 2017, 7: 678. |
| [46] |
STAKOS D A, KAMBAS K, KONSTANTINIDIS T, et al. Expression of functional tissue factor by neutrophil extracellular traps in culprit artery of acute myocardial infarction[J]. European Heart Journal, 2015, 36(22): 1405-1414. DOI:10.1093/eurheartj/ehv007 |
| [47] |
PARK S Y, SHRESTHA S, YOUN Y J, et al. Autophagy primes neutrophils for neutrophil extracellular trap formation during sepsis[J]. American Journal of Respiratory and Critical Care Medicine, 2017, 196(5): 577-589. DOI:10.1164/rccm.201603-0596OC |
| [48] |
MAO J Y, ZHANG J H, CHENG W, et al. Effects of neutrophil extracellular traps in patients with septic coagulopathy and their interaction with autophagy[J]. Frontiers in Immunology, 2021, 12: 757041. DOI:10.3389/fimmu.2021.757041 |
| [49] |
TEIMOURIAN S, MOGHANLOO E. Role of PTEN in neutrophil extracellular trap formation[J]. Molecular Immunology, 2015, 66(2): 319-324. DOI:10.1016/j.molimm.2015.03.251 |
| [50] |
GERMIC N, STOJKOV D, OBERSON K, et al. Neither eosinophils nor neutrophils require ATG5-dependent autophagy for extracellular DNA trap formation[J]. Immunology, 2017, 152(3): 517-525. DOI:10.1111/imm.12790 |
| [51] |
REMIJSEN Q, VANDEN BERGHE T, WIRAWAN E, et al. Neutrophil extracellular trap cell death requires both autophagy and superoxide generation[J]. Cell Research, 2011, 21(2): 290-304. DOI:10.1038/cr.2010.150 |
| [52] |
WANG Y P, GAO W Q, SHI X Y, et al. Chemotherapy drugs induce pyroptosis through Caspase-3 cleavage of a gasdermin[J]. Nature, 2017, 547(7661): 99-103. DOI:10.1038/nature22393 |
| [53] |
LISTON A, MASTERS S L. Homeostasis-altering molecular processes as mechanisms of inflammasome activation[J]. Nature Reviews Immunology, 2017, 17(3): 208-214. DOI:10.1038/nri.2016.151 |
| [54] |
BURDETTE B E, ESPARZA A N, ZHU H, et al. Gasdermin D in pyroptosis[J]. Acta Pharmaceutica Sinica B, 2021, 11(9): 2768-2782. DOI:10.1016/j.apsb.2021.02.006 |
| [55] |
ZHANG H, CHEN Z, ZHOU J, et al. NAT10 regulates neutrophil pyroptosis in sepsis via acetylating ULK1 RNA and activating STING pathway[J]. Communications Biology, 2022, 5(1): 916. DOI:10.1038/s42003-022-03868-x |
| [56] |
GENTILE L F, CUENCA A L, CUENCA A G, et al. Improved emergency myelopoiesis and survival in neonatal sepsis by Caspase-1/ 11 ablation[J]. Immunology, 2015, 145(2): 300-311. DOI:10.1111/imm.12450 |
| [57] |
KOVACS S B, OH C, MALTEZ V I, et al. Neutrophil caspase-11 is essential to defend against a cytosol-invasive bacterium[J]. Cell Reports, 2020, 32(4): 107967. DOI:10.1016/j.celrep.2020.107967 |
| [58] |
SILVA C M S, WANDERLEY C W S, VERAS F P, et al. Gasdermin D inhibition prevents multiple organ dysfunction during sepsis by blocking NET formation[J]. Blood, 2021, 138(25): 2702-2713. DOI:10.1182/blood.2021011525 |
| [59] |
CAUTION K, YOUNG N, ROBLEDO-AVILA F, et al. Caspase-11 mediates neutrophil chemotaxis and extracellular trap formation during acute gouty arthritis through alteration of cofilin phosphorylation[J]. Frontiers in Immunology, 2019, 10: 2519. DOI:10.3389/fimmu.2019.02519 |
| [60] |
XIE J, ZHU C L, WAN X J, et al. GSDMD-mediated NETosis promotes the development of acute respiratory distress syndrome[J]. European Journal of Immunology, 2023, 53(1): 2250011. DOI:10.1002/eji.202250011 |
| [61] |
LI C Y, WANG P, LI M, et al. The current evidence for the treatment of sepsis with Xuebijing Injection: Bioactive constituents, findings of clinical studies and potential mechanisms[J]. Journal of Ethnopharmacology, 2021, 265: 113301. DOI:10.1016/j.jep.2020.113301 |
| [62] |
CHEN W J, ZHAO J J, MU D, et al. Pyroptosis mediates neutrophil extracellular trap formation during bacterial infection in zebrafish[J]. Journal of Immunology, 2021, 206(8): 1913-1922. DOI:10.4049/jimmunol.2001335 |
| [63] |
SHANG T, ZHANG Z S, WANG X T, et al. Xuebijing Injection inhibited neutrophil extracellular traps to reverse lung injury in sepsis mice via reducing Gasdermin D[J]. Frontiers in Pharmacology, 2022, 13: 1054176. DOI:10.3389/fphar.2022.1054176 |
| [64] |
LV J, GUO X X, ZHAO H Y, et al. Xuebijing administration alleviates pulmonary endothelial inflammation and coagulation dysregulation in the early phase of sepsis in rats[J]. Journal of Clinical Medicine, 2022, 11(22): 6696. DOI:10.3390/jcm11226696 |
| [65] |
WANG Y P, GUO Y, WEN P S, et al. Three ingredients of safflower alleviate acute lung injury and inhibit NET release induced by lipopolysaccharide[J]. Mediators of Inflammation, 2020, 2020(1): 2720369. |
| [66] |
ZHOU M G, LIU Y M, QIN H L, et al. Xuanfei Baidu Decoction regulates NETs formation via CXCL2 / CXCR2 signaling pathway that is involved in acute lung injury[J]. Biomedicine & Pharmaco-therapy, 2023, 161: 114530. |
| [67] |
曾仪晨, 马雨雯, 曾榆淞, 等. 四妙勇安汤加减化裁治疗脓毒症的研究进展及其应用开发思路与方法[J]. 中草药, 2025, 56(1): 330-339. |
| [68] |
JIE S S, SUN H J, LIU J X, et al. Simiao Yong'an decoction ameliorates murine collagen-induced arthritis by modulating neutrophil activities: An in vitro and in vivo study[J]. Journal of Ethnopharmacology, 2023, 305: 116119. DOI:10.1016/j.jep.2022.116119 |
| [69] |
YANG C X, SONG C L, LIU Y T, et al. Re-Du-Ning injection ameliorates LPS-induced lung injury through inhibiting neutrophil extracellular traps formation[J]. Phytomedicine, 2021, 90: 153635. DOI:10.1016/j.phymed.2021.153635 |
| [70] |
ZHANG H Y, SONG X J, GE S N, et al. Zixue Powder attenuates septic thrombosis via reducing neutrophil extracellular trap through blocking platelet STING activation[J]. Journal of Ethnopharmacology, 2024, 331: 118337. DOI:10.1016/j.jep.2024.118337 |
| [71] |
LEI B, MU J W, XU G H, et al. Jing-Yin-Gu-Biao formula protects mice from postinfluenza Staphylococcus aureus infection by ameliorating acute lung injury and improving hypercoagulable state via inhibiting NETosis[J]. Frontiers in Immunology, 2025, 16: 1567522. DOI:10.3389/fimmu.2025.1567522 |
| [72] |
YANG K, WU B, WEI W, et al. Curdione ameliorates sepsis-induced lung injury by inhibiting platelet-mediated neutrophil extracellular trap formation[J]. International Immunopharmacology, 2023, 118: 110082. DOI:10.1016/j.intimp.2023.110082 |
| [73] |
XIE S H, DAI J H, ZHANG Y K, et al. Effects of glycyrrhizin on the expression of HMGB1 and inflammatory factors in macrophages and blood of neonatal rats with sepsis[J]. Panminerva Medica, 2020. |
| [74] |
ZHAO F, FANG Y, DENG S X, et al. Glycyrrhizin protects rats from sepsis by blocking HMGB1 signaling[J]. BioMed Research International, 2017, 2017(1): 9719647. |
| [75] |
GU J R, RAN X F, DENG J, et al. Glycyrrhizin alleviates sepsis-induced acute respiratory distress syndrome via suppressing of HMGB1/TLR9 pathways and neutrophils extracellular traps formation[J]. International Immunopharmacology, 2022, 108: 108730. DOI:10.1016/j.intimp.2022.108730 |
| [76] |
HUANG Y, WU Y, YIN H X, et al. Senkyunolide Ⅰ: A review of its phytochemistry, pharmacology, pharmacokinetics, and drug-likeness[J]. Molecules, 2023, 28(8): 3636. DOI:10.3390/molecules28083636 |
| [77] |
ZHA Y F, XIE J, DING P, et al. Senkyunolide I protect against lung injury via inhibiting formation of neutrophil extracellular trap in a murine model of cecal ligation and puncture[J]. International Immunopharmacology, 2021, 99: 107922. DOI:10.1016/j.intimp.2021.107922 |
| [78] |
HE W J, XI Q, CUI H T, et al. Forsythiaside B ameliorates coagulopathies in a rat model of sepsis through inhibition of the formation of PAD4-dependent neutrophil extracellular traps[J]. Frontiers in Pharmacology, 2022, 13: 1022985. DOI:10.3389/fphar.2022.1022985 |
| [79] |
LU Q Y, JIANG H X, ZHU Q Q, et al. Tetrandrine ameliorates rheumatoid arthritis in mice by alleviating neutrophil activities[J]. Evidence-Based Complementary and Alternative Medicine, 2022, 2022(1): 8589121. |
| [80] |
GAO F, LIU X, SHEN Z Y, et al. Andrographolide sulfonate attenuates acute lung injury by reducing expression of myeloperoxidase and neutrophil-derived proteases in mice[J]. Frontiers in Physiology, 2018, 9: 939. DOI:10.3389/fphys.2018.00939 |
| [81] |
HUANG G R, YUAN K, ZHU Q Q, et al. Triptolide inhibits the inflammatory activities of neutrophils to ameliorate chronic arthritis[J]. Molecular Immunology, 2018, 101: 210-220. DOI:10.1016/j.molimm.2018.06.012 |
| [82] |
GUAN H Y, XIE L F, JI Z Z, et al. Triptolide inhibits neutrophil extracellular trap formation[J]. Annals of Translational Medicine, 2021, 9(17): 1384. DOI:10.21037/atm-21-3522 |
| [83] |
YANG H L, QIAN H, LIU B, et al. Triptolide dose-dependently improves LPS-induced alveolar hypercoagulation and fibrinolysis inhibition through NF-κB inactivation in ARDS mice[J]. Biomedicine & Pharmacotherapy, 2021, 139: 111569. |
| [84] |
WU Q, CHEN Y C, ZHU B Q, et al. Bletilla striata polysaccharides alleviate ARDS by inhibiting NETs-induced pyroptosis in pulmonary alveolar macrophage through the PAD4 pathway[J]. International Journal of Biological Macromolecules, 2025, 305: 141026. DOI:10.1016/j.ijbiomac.2025.141026 |
| [85] |
XIONG Y, HALIMA M, CHE X Y, et al. Steamed Panax notoginseng and its saponins inhibit the migration and induce the apoptosis of neutrophils in a zebrafish tail-fin amputation model[J]. Frontiers in Pharmacology, 2022, 13: 946900. DOI:10.3389/fphar.2022.946900 |
| [86] |
PARK H H, KIM H, LEE H S, et al. PEGylated nanoparticle albumin-bound steroidal ginsenoside derivatives ameliorate SARSCoV-2-mediated hyper-inflammatory responses[J]. Biomaterials, 2021, 273: 120827. DOI:10.1016/j.biomaterials.2021.120827 |
| [87] |
YANG S C, CHEN P J, CHANG S H, et al. Luteolin attenuates neutrophilic oxidative stress and inflammatory arthritis by inhibiting Raf1 activity[J]. Biochemical Pharmacology, 2018, 154: 384-396. DOI:10.1016/j.bcp.2018.06.003 |
| [88] |
ZHANG Z H, YANG H X, JIN Q, et al. Luteolin attenuates hepatic injury in septic mice by regulating P2X7R-based HMGB1 release[J]. Food & Function, 2021, 12(21): 10714-10727. |