 2026, Vol. 43
2026, Vol. 43文章信息
- 卢照平, 何健东, 谭志超, 等.
- LU Zhaoping, HE Jiandong, TAN Zhichao, et al.
- 姜黄素调节Notch1/Hes1/Prdx1通路对IL-1β诱导的软骨细胞炎性损伤的影响
- The effect of curcumin regulating the Notch1/Hes1/Prdx1 pathway on IL-1β-induced inflammatory injury of chondrocytes
- 天津中医药, 2026, 43(6): 749-755
- Tianjin Journal of Traditional Chinese Medicine, 2026, 43(6): 749-755
- http://dx.doi.org/10.11656/j.issn.1672-1519.2026.06.12
-
文章历史
- 收稿日期: 2026-01-12

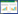



骨关节炎(OA)是全球最常见的关节疾病之一,其特征是关节软骨逐渐退化、继发性间歇性滑膜炎和骨重建,导致关节功能障碍和身体残疾,并伴有长期难以忍受的疼痛[1]。越来越多的证据表明,炎症反应在OA进展中至关重要,其中炎症细胞因子发挥关键作用[2]。作为炎症细胞因子家族的一员,白细胞介素-1β(IL-1β)在OA患者滑膜液和软骨组织中表达升高,据报道IL-1β通过调节软骨细胞中的细胞因子来打破体内平衡,从而参与OA的病理生理过程,IL-1β诱导的软骨细胞常用于体外模拟OA关节软骨损伤[3]。因此,探讨IL-1β诱导的软骨细胞炎性损伤的分子机制具有重要意义。Notch1信号通路是一种近分泌通路,在该通路中,受体胞内结构域在配体结合后被切割;随后,该结构域可转运至细胞核内,激活靶基因Hes1的表达,Hes1是一个转录抑制因子,通过结合到Prdx1基因启动子区的特定序列,直接抑制Prdx1的转录,研究表明该信号通路的激活可以促进OA疾病进展[4]。目前,OA的主要治疗方法是减少关节的承重和过度活动范围,从而延缓病情的发展。目前常用的抗炎和止痛药物可以减轻或控制症状,但不能长期服用[5]。因此,寻找治疗OA的新药物具有重要意义。姜黄素(Cur)是一种从姜黄根茎中提取的多酚类色素,因其具有良好的特性而被广泛用于缓解炎症和退行性疾病,体内、外研究已证明Cur在治疗OA方面具有较高疗效[6]。相关研究显示,Cur可以抑制Notch信号通路对T细胞急性淋巴细胞白血病具有良好的治疗效果[7]。然而,Cur调节Notch1/Hes1/Prdx1通路对IL-1β诱导的软骨细胞炎性损伤的影响,目前尚未明确。基于此,本研究对此展开探讨,旨在为优化OA的临床治疗效果提供实验依据与理论支持。
1 材料与方法 1.1 材料及仪器人正常软骨细胞C28/I2细胞(货号:SHC1968,南京安元生物公司);Cur(货号:C1386,嘉兴硕玺生物公司);多聚甲醛(货号:P804536,北京萘析生化科技公司);Notch激活剂Jagged 1(货号:TP310516,无锡傲锐东源生物公司);MTS细胞增殖试剂盒(货号:HL10043,上海哈灵生物公司);白细胞介素(IL)-6、IL-8、肿瘤坏死因子-α(TNF-α)、基质金属蛋白酶(MMP)-3、MMP-13酶联免疫吸附测定(ELISA)试剂盒(货号:KJ-0049、KJ-0088、KJ-0122、KJ-0108、KJ-0070,江苏科晶生物公司);β-actin、Ⅱ型胶原(ColⅡ)、聚集蛋白聚糖(Aggrecan)、增殖细胞核抗原(Ki-67)、微染色体维持复合体组分7(MCM)7、Notch1、Hes1、Prdx1、BCL2拮抗剂(Bak)、存活蛋白(survivin)、HRP(货号:ab8227、ab188570、ab186414、ab92742、ab179904、ab52627、ab71559、ab109498、ab69404、ab469、ab205718,英国Abcam公司);流式细胞仪(型号:Lymacyte,上海诺亚源医学科技公司);倒置显微镜(型号:HCS321Gi,上海恒商精密仪器公司)。
1.2 方法 1.2.1 细胞培养与分组C28/I2细胞随机分为对照(Ctrl)组(正常培养)、IL-1β组(10 ng/mL IL-1β[8])、Cur低剂量(L-Cur)、Cur中剂量(M-Cur)、Cur高剂量(H-Cur)组(IL-1β诱导+10、20、40 μmol/L Cur[9])、Cur+锯齿蛋白1(Jagged 1)组(IL-1β诱导+40 μmol/L Cur+0.5 μg/mL Notch激活剂Jagged 1[10])。
1.2.2 MTS法和5-乙炔基-2'-脱氧尿苷(EdU)染色检测C28/I2细胞增殖MTS法:以每孔5×103个的密度将C28/I2细胞接种于96孔培养板培养24 h,每孔加入20 μL MTS试剂孵育4 h,使用酶标仪在490 nm波长下测定各孔的吸光度(OD值)。
EdU染色:将C28/I2细胞以5×104个/孔的密度接种在24孔板中,细胞核用DAPI染色,用荧光显微镜观察并检测。
1.2.3 流式细胞仪检测C28/I2细胞凋亡将C28/I2细胞以每孔2.0×105个细胞的密度接种到6孔板中,在黑暗中于室温下将细胞与FITC标记Annexin V和PI一起染色15 min,使用流式细胞仪分析。
1.2.4 2',7'-二氯荧光黄双乙酸盐(DCFH-DA)荧光探针检测C28/I2细胞中ROS水平收集C28/I2细胞在37 ℃、避光条件下,用终浓度为10 μmol的DCFH-DA探针负载细胞,用PBS清洗去除未内化的探针,于显微镜下观察。
1.2.5 免疫荧光检测C28/I2细胞ColⅡ、Aggrecan表达制备C28/I2细胞爬片,加1 mL 4 %多聚甲醛室温固定15 min,加1 mL 0.1% Triton X-100孵育10 min,加1 mL 5 % BSA孵育60 min,加入ColⅡ(1∶100)、Aggrecan(1∶500)一抗4℃孵育过夜,加入二抗(1∶2 000)避光孵育60 min,加DAPI染液室温避光孵育5 min,进行封片与成像,在Image J中,手动或自动划定每个细胞的感兴趣区域(ROI),软件会输出每个ROI的积分荧光密度和区域面积,计算平均荧光强度=积分荧光密度/区域面积。
1.2.6 ELISA试剂盒检测C28/I2细胞中IL-6、IL-8、TNF-α、MMP-3、MMP-13表达收集C28/I2细胞以3 500 r/min(半径10 cm)离心10 min取上清,将相应ELISA试剂盒试剂在室温下放置30 min,按所需标准依次添加样本和试剂,在37 ℃下30 min,向孔中加入终止液并反应5 min,通过酶标仪测定450 nm处的OD值,并根据标准曲线计算浓度。
1.2.7 Western Blot检测C28/I2细胞中Ki-67、MCM7、Notch1、Hes1、Prdx1、Bak、survivin蛋白表达在裂解缓冲液中裂解C28/I2细胞(临用前按RIPA∶PMSF=100∶1加入PMSF),在4 ℃下以12 000×g速度离心20 min,使用Lowry法测定蛋白质浓度,通过10 % SDS-PAGE分离,转移到PVDF膜上,并封存2 h,在4 ℃下与β-actin(1∶1 000)、Ki-67(1∶5 000)、MCM7(1∶1 000)、Notch1(1∶1 000)、Hes1(1∶200)、Prdx1(1∶10 000)、Bak(1∶1 000)、survivin(1∶5 000)一抗孵育过夜,与二抗(1∶2 000)孵育1 h,使用增强ECL对蛋白质进行显色。
1.3 统计学分析采用SPSS 23.0软件进行数据分析,符合正态分布的计量资料,以均数±标准差(x±s)表示,单因素方差分析用于多组间比较,进一步两两比较行SNK-q检验,P<0.05表示差异有统计学意义。
2 结果 2.1 Cur对C28/I2细胞增殖的影响IL-1β组OD490值、EdU阳性细胞率、Ki-67、MCM7低于Ctrl组(P<0.05);L-Cur组、M-Cur组、H-Cur组OD490值、EdU阳性细胞率、Ki-67、MCM7高于IL-1β组(P<0.05);Cur+Jagged 1组OD490值、EdU阳性细胞率、Ki-67、MCM7低于H-Cur组(P<0.05)。见图 1、4、表 1。

|
| 图 1 EdU染色检测C28/I2细胞增殖(×200) Fig. 1 EdU staining for detecting the proliferation of C28/I2 cells(×200) |
| 组别 | n | OD490值 | EdU阳性细胞率(%) | Ki-67 | MCM7 |
| Ctrl组 | 6 | 0.89±0.12 | 48.35±4.96 | 1.12±0.18 | 0.73±0.11 |
| IL-1β组 | 6 | 0.23±0.07* | 12.24±1.86* | 0.47±0.12* | 0.22±0.06* |
| L-Cur组 | 6 | 0.41±0.09# | 21.17±2.65# | 0.69±0.14# | 0.39±0.08# |
| M-Cur组 | 6 | 0.63±0.10#△ | 30.56±3.82#△ | 0.86±0.16#△ | 0.56±0.09#△ |
| H-Cur组 | 6 | 0.81±0.11#△▲ | 43.82±4.73#△▲ | 1.04±0.17#△▲ | 0.67±0.10#△▲ |
| Cur+Jagged 1组 | 6 | 0.35±0.08& | 19.78±2.14& | 0.58±0.13& | 0.34±0.07& |
| 注:与Ctrl组比较,*P<0.05;与IL-1β组比较,#P<0.05;与L-Cur组比较,△P<0.05;与M-Cur组比较,▲P<0.05;与H-Cur组比较,&P<0.05。 | |||||
IL-1β组凋亡率、Bak高于Ctrl组,survivin低于Ctrl组(P<0.05);L-Cur组、M-Cur组、H-Cur组凋亡率、Bak低于IL-1β组,survivin高于IL-1β组(P<0.05);Cur+Jagged 1组凋亡率、Bak高于H-Cur组,survivin低于H-Cur组(P<0.05)。见图 2、4、表 2。

|
| 图 2 流式细胞仪检测C28/I2细胞凋亡 Fig. 2 Flow cytometry detection of apoptosis in C28/I2 cells |
| 组别 | n | 凋亡率(%) | Bak | survivin |
| Ctrl组 | 6 | 1.17±0.21 | 0.28±0.08 | 0.93±0.15 |
| IL-1β组 | 6 | 37.39±4.16* | 0.96±0.15* | 0.31±0.09* |
| L-Cur组 | 6 | 25.82±2.73# | 0.75±0.13# | 0.49±0.11# |
| M-Cur组 | 6 | 17.56±2.08#△ | 0.54±0.12#△ | 0.67±0.13#△ |
| H-Cur组 | 6 | 9.16±1.16#△▲ | 0.32±0.09#△▲ | 0.83±0.14#△▲ |
| Cur+Jagged 1组 | 6 | 32.24±3.59& | 0.83±0.14& | 0.42±0.10& |
| 注:与Ctrl组比较,*P<0.05;与IL-1β组比较,#P<0.05;与L-Cur组比较,△P<0.05;与M-Cur组比较,▲P<0.05;与H-Cur组比较,&P<0.05。 | ||||
IL-1β组ROS荧光强度高于Ctrl组(P<0.05);L-Cur组、M-Cur组、H-Cur组ROS荧光强度低于IL-1β组(P<0.05);Cur+Jagged 1组ROS荧光强度高于H-Cur组(P<0.05)。见图 3、表 3。

|
| 图 3 DCFH-DA荧光探针检测C28/I2细胞中ROS水平(×200) Fig. 3 Detection of ROS levels in C28/I2 cells using DCFH-DA fluorescent probe(×200) |
| 组别 | n | ROS荧光强度(RFU) |
| Ctrl组 | 6 | 104.54±27.32 |
| IL-1β组 | 6 | 396.68±41.06* |
| L-Cur组 | 6 | 306.51±35.59# |
| M-Cur组 | 6 | 234.72±33.24#△ |
| H-Cur组 | 6 | 136.24±29.08#△▲ |
| Cur+Jagged 1组 | 6 | 348.47±37.72& |
| 注:与Ctrl组比较,*P<0.05;与IL-1β组比较,#P<0.05;与L-Cur组比较,△P<0.05;与M-Cur组比较,▲P<0.05;与H-Cur组比较,&P<0.05。 | ||
IL-1β组MMP-3、MMP-13高于Ctrl组,ColⅡ荧光强度、Aggrecan荧光强度低于Ctrl组(P<0.05);L-Cur组、M-Cur组、H-Cur组MMP-3、MMP-13低于IL-1β组,ColⅡ荧光强度、Aggrecan荧光强度高于IL-1β组(P<0.05);Cur+Jagged 1组MMP-3、MMP-13高于H-Cur组,ColⅡ荧光强度、Aggrecan荧光强度低于H-Cur组(P<0.05)。见图 4、表 4。

|
| 图 4 免疫荧光检测C28/I2细胞ColⅡ、Aggrecan表达(×200) Fig. 4 Immunofluorescence detection of ColⅡ and Aggrecan expression in C28/I2 cells(×200) |
| 组别 | n | ColⅡ荧光强度(RFU) | Aggrecan荧光强度(RFU) | MMP-3(pg/mL) | MMP-13(pg/mL) |
| Ctrl组 | 6 | 96.54±9.87 | 82.31±8.35 | 34.19±4.15 | 9.28±1.48 |
| IL-1β组 | 6 | 38.16±5.46* | 26.14±4.29* | 96.24±9.86* | 35.36±4.05* |
| L-Cur组 | 6 | 53.29±7.15# | 43.26±6.41# | 75.47±7.95# | 28.43±2.97# |
| M-Cur组 | 6 | 71.62±8.26#△ | 61.07±7.38#△ | 53.13±6.52#△ | 21.12±2.42#△ |
| H-Cur组 | 6 | 92.37±9.43#△▲ | 78.59±8.14#△▲ | 39.87±5.34#△▲ | 14.06±1.96#△▲ |
| Cur+Jagged 1组 | 6 | 41.08±6.32& | 31.48±5.27& | 83.49±8.71& | 31.29±3.47& |
| 注:与Ctrl组比较,*P<0.05;与IL-1β组比较,#P<0.05;与L-Cur组比较,△P<0.05;与M-Cur组比较,▲P<0.05;与H-Cur组比较,&P<0.05。 | |||||
IL-1β组IL-6、IL-8、TNF-α表达高于Ctrl组(P<0.05);L-Cur组、M-Cur组、H-Cur组IL-6、IL-8、TNF-α表达低于IL-1β组(P<0.05);Cur+Jagged 1组IL-6、IL-8、TNF-α表达高于H-Cur组(P<0.05)。见表 5。
| pg/mL | |||||||||||||||||||||||||||||
| 组别 | n | IL-6 | IL-8 | TNF-α | |||||||||||||||||||||||||
| Ctrl组 | 6 | 53.19±9.17 | 18.32±2.09 | 3.12±0.86 | |||||||||||||||||||||||||
| IL-1β组 | 6 | 154.28±15.83* | 56.49±5.82* | 19.36±2.73* | |||||||||||||||||||||||||
| L-Cur组 | 6 | 129.35±13.27# | 43.27±4.76# | 15.27±1.97# | |||||||||||||||||||||||||
| M-Cur组 | 6 | 98.72±11.05#△ | 34.16±3.93#△ | 11.48±1.74#△ | |||||||||||||||||||||||||
| H-Cur组 | 6 | 62.64±10.32#△▲ | 21.53±2.87#△▲ | 7.51±1.25#△▲ | |||||||||||||||||||||||||
| Cur+Jagged 1组 | 6 | 137.59±14.32& | 49.07±5.03& | 17.74±2.28& | |||||||||||||||||||||||||
| 注:与Ctrl组比较,*P<0.05;与IL-1β组比较,#P<0.05;与L-Cur组比较,△P<0.05;与M-Cur组比较,▲P<0.05;与H-Cur组比较,&P<0.05。 | |||||||||||||||||||||||||||||
IL-1β组Notch1、Hes1、Prdx1表达高于Ctrl组(P<0.05);L-Cur组、M-Cur组、H-Cur组Notch1、Hes1、Prdx1表达低于IL-1β组(P<0.05);Cur+Jagged 1组Notch1、Hes1、Prdx1表达高于H-Cur组(P<0.05)。见图 5、表 6。

|
| 图 5 Western Blot检测C28/I2细胞中Ki-67、MCM7、Notch1、Hes1、Prdx1、Bak、survivin蛋白表达 Fig. 5 Western Blot analysis of the expression of Ki-67, MCM7, Notch1, Hes1, Prdx1, Bak and survivin proteins in C28/I2 cells |
| 组别 | n | Notch1 | Hes1 | Prdx1 |
| Ctrl组 | 6 | 0.58±0.12 | 0.23±0.06 | 0.36±0.11 |
| IL-1β组 | 6 | 1.47±0.18* | 0.98±0.12* | 1.05±0.16* |
| L-Cur组 | 6 | 1.13±0.15# | 0.76±0.10# | 0.83±0.04# |
| M-Cur组 | 6 | 0.89±0.14#△ | 0.54±0.09#△ | 0.59±0.13#△ |
| H-Cur组 | 6 | 0.65±0.13#△▲ | 0.31±0.07#△▲ | 0.41±0.12#△▲ |
| Cur+Jagged 1组 | 6 | 1.29±0.17& | 0.85±0.11& | 0.98±0.15& |
| 注:与Ctrl组比较,*P<0.05;与IL-1β组比较,#P<0.05;与L-Cur组比较,△P<0.05;与M-Cur组比较,▲P<0.05;与H-Cur组比较,&P<0.05。 | ||||
软骨退化是OA进展的主要原因,各种软骨细胞损伤会直接加速这一过程[11]。IL-1β是一种重要的炎症因子,参与OA的起始和早期进展,据报道,暴露于IL-1β的软骨细胞会释放炎症因子和介质,如IL-6、IL-8、TNF-α,直接诱导软骨细胞内ROS的大量生成,还可通过正反馈循环进一步放大IL-1β的促炎效应,加重OA的发病机制[12]。炎症细胞因子通过抑制软骨细胞的合成代谢活动(ColⅡ、Aggrecan表达降低)对软骨造成破坏性影响,并激活软骨细胞产生基质金属蛋白酶(MMPs),尤其是MMP-13、MMP-3,诱导软骨退化[13]。除了破坏性作用外,炎症性细胞因子还倾向于诱导软骨细胞的凋亡,进一步促进OA疾病进展[14]。Bak、survivin分别可作为细胞凋亡和抑制凋亡的蛋白标志物;Ki-67、MCM7是常用作细胞增殖蛋白标志物[15]。本研究采用IL-1β诱导的软骨细胞结果发现,细胞增殖能力明显下降,凋亡增多,促炎因子表达和细胞外基质降解增加,提示IL-1β通过促进软骨细胞内炎症反应和氧化应激,引起软骨细胞骨代谢失衡、细胞凋亡,促进OA进展。
在经典的Notch信号通路中,NICD进入细胞核并与共激活因子MAML复合物一起形成RBP-J,最终激活下游效应物(如Hes家族蛋白)的表达,该通路的异常激活在OA进展中发挥重要作用[16]。Prdx1是一种抗氧化剂,在应激条件下,尤其是各种炎症反应期间,Prdx1会被大量诱导表达,由于Prdx1在细胞内能够清除过氧化物和过氧亚硝酸盐,在炎症性疾病中发挥保护作用[17]。肖惟丹等[18]发现,巴戟天通过抑制Notch1/Hes1/Prdx1通路,减少骨质疏松大鼠骨量流失,改善骨组织生物力学。鲁林等[19]发现,骨碎补总黄酮通过抑制Notch1/Hes1/Prdx1通路,抑制氧化应激,调控骨稳态,缓解骨质疏松。以上研究表明抑制Notch1/Hes1/Prdx1通路可以抑制氧化应激、炎症反应,进而减轻骨相关疾病进展。本研究发现,在IL-1β诱导的软骨细胞中Notch1、Hes1、Prdx1蛋白表达升高,提示IL-1β可激活氧化应激,促进Notch1/Hes1信号通路上调,而氧化应激同时驱动Prdx1转录表达以缓解氧化损伤,Hes1对Prdx1的转录抑制作用被氧化应激效应所拮抗,最终表现为Prdx1高表达[16-17]。在IL-1β的持续刺激下,其抗氧化能力最终会被耗竭,进一步促进炎症反应,进而引发软骨细胞不可逆凋亡与骨代谢失衡,推动OA进展[4]。
Cur具有丰富的生物活性,包括抗炎、抗氧化和抗凋亡特性,能够调节多种分子靶点和信号通路,广泛应用于治疗OA等多种关节炎疾病[20]。Jin等[21]发现,Cur通过调节促进AMPK/PINK1/Parkin介导的线粒体自噬,在体外抑制IL-1β诱导的软骨细胞炎性损伤,在体内对OA大鼠发挥软骨保护作用。Wang等[22]发现,Cur抑制P38MAPK磷酸化来减轻软骨细胞的炎症反应,调节氧化应激并维持线粒体功能,在延缓OA大鼠软骨基质分解的同时阻止软骨细胞凋亡。Chen等[23]发现,Cur通过激活自噬并抑制NF-κB信号通路,改善IL-1β诱导的大鼠原代关节软骨细胞凋亡。Li等[24]发现,Cur通过抑制Bmp2/Smad5/Runx2通路,减轻IL-1β诱导的软骨细胞氧化应激、炎症反应(TNF-α、IL-6表达)和细胞凋亡,并增加细胞外基质的分泌,从而对受损的软骨细胞发挥保护作用。上述发现说明,Cur可以激活线粒体自噬,抑制炎症反应、氧化应激、细胞凋亡、细胞外基质降解,进而缓解OA进展。本研究发现,Cur可以抑制Notch1/Hes1/Prdx1通路,抑制IL-1β诱导的炎症反应、氧化应激、细胞凋亡、细胞外基质降解,而Notch激活剂则逆转Cur的作用,提示Cur可以抑制IL-1β诱导的软骨细胞炎性损伤,其机制可能是通过抑制Notch1/Hes1/Prdx1通路实现的。
综上所述,Cur通过抑制Notch1/Hes1/Prdx1通路,减轻IL-1β诱导的软骨细胞炎性损伤。然而,本研究通常采用IL-1β诱导的体外软骨细胞模型,但模型难以完全模拟OA关节腔内复杂的炎症环境,且缺乏动物实验和临床验证,后续会补充动物和临床实验,继续研究。
| [1] |
LIU C, CHEN Y. Ketorolac tromethamine alleviates IL-1β-induced chondrocyte injury by inhibiting COX-2 expression[J]. Experimental and Therapeutic Medicine, 2022, 23(5): 337. DOI:10.3892/etm.2022.11267 |
| [2] |
JIANG J, CAI M. Cardamonin inhibited IL-1β induced injury by inhibition of NLRP3 inflammasome via activating Nrf2/NQO-1 signaling pathway in chondrocyte[J]. Journal of Microbiology and Biotechnology, 2021, 31(6): 794-802. DOI:10.4014/jmb.2103.03057 |
| [3] |
OU D, LIU S, TONG C, et al. LIM mineralization protein-1 inhibits IL-1β-induced human chondrocytes injury by altering the NF-κB and MAPK/JNK pathways[J]. Experimental and Therapeutic Medicine, 2022, 23(1): 61. |
| [4] |
FAZIO A, DI MARTINO A, BRUNELLO M, et al. The involvement of signaling pathways in the pathogenesis of osteoarthritis: An update[J]. Journal of Orthopaedic Translation, 2024, 47(1): 116-124. |
| [5] |
LUO X, LIAO H, PENG J, et al. Salidroside protects chondrocytes against IL-1β-induced injury and alleviates osteoarthritis progression by activating the Nrf2 pathway[J]. Discovery Medicine, 2024, 36(181): 266-277. DOI:10.24976/Discov.Med.202436181.25 |
| [6] |
ZHAO J, LIANG G, ZHOU G, et al. Efficacy and safety of curcumin therapy for knee osteoarthritis: A Bayesian network Meta-analysis[J]. Journal of Ethnopharmacology, 2024, 321(1): 117493-117504. |
| [7] |
ZHDANOVSKAYA N, LAZZARI S, CAPRIOGLIO D, et al. Identification of a novel curcumin derivative influencing notch pathway and DNA damage as a potential therapeutic agent in T-ALL[J]. Cancers, 2022, 14(23): 5772. DOI:10.3390/cancers14235772 |
| [8] |
张应鹏, 王世金, 亓峰. 白术内酯Ⅰ对IL-1β诱导的关节软骨细胞炎性损伤的影响[J]. 中国骨质疏松杂志, 2024, 30(8): 1152-1156. |
| [9] |
李杨, 路坦, 庄全魁, 等. 基于铁死亡途径探究姜黄素对骨关节炎模型大鼠的骨保护作用[J]. 河北医学, 2025, 31(4): 591-597. |
| [10] |
蒋智永, 席庆春, 李旋, 等. 黄芪甲苷调节Notch信号通路对瘢痕疙瘩成纤维细胞增殖和迁移的影响[J]. 中国细胞生物学学报, 2025, 47(4): 832-840. |
| [11] |
CAO S, WEI Y, XIONG A, et al. Paeonol inhibits ACSL4 to protect chondrocytes from ferroptosis and ameliorates osteoarthritis progression[J]. Journal of Orthopaedic Translation, 2024, 50(1): 1-13. |
| [12] |
HUANG Y, CHEN D, YAN Z, et al. LncRNA MEG3 protects chondrocytes from IL-1β-induced inflammation via regulating miR-9-5p/KLF4 Axis[J]. Frontiers in Physiology, 2021, 12(1): 617654-617663. |
| [13] |
WU J, QIAN Y, CHEN C, et al. Hesperetin exhibits anti-inflammatory effects on chondrocytes via the AMPK pathway to attenuate anterior cruciate ligament transection-induced osteoarthritis[J]. Frontiers in Pharmacology, 2021, 12(1): 735087-735097. |
| [14] |
JOUNG S, YOON D S, CHO S, et al. Downregulation of MicroRNA-495 alleviates IL-1β Responses among chondrocytes by preventing SOX9 reduction[J]. Yonsei Medical Journal, 2021, 62(7): 650-659. DOI:10.3349/ymj.2021.62.7.650 |
| [15] |
GUO Y, LI R, DANG X. S100A10 regulates tumor necrosis factor alpha-induced apoptosis in chondrocytes via the reactive oxygen species/nuclear factor-kappa B pathway[J]. Biotechnology and Applied Biochemistry, 2022, 69(6): 2284-2295. DOI:10.1002/bab.2285 |
| [16] |
FENG J, ZHANG Q, PU F, et al. Signalling interaction between β-catenin and other signalling molecules during osteoarthritis development[J]. Cell Proliferation, 2024, 57(6): 13600-13614. DOI:10.1111/cpr.13600 |
| [17] |
KANG J H, LEE H Y, KIM N Y, et al. Extracellular Prdx1 mediates bacterial infection and inflammatory bone diseases[J]. Life Sciences, 2023, 333(1): 122140. |
| [18] |
肖惟丹, 熊政利, 李俊. 巴戟天调控Notch1/Hes1/Prdx1通路对绝经后骨质疏松大鼠骨质量、生物力学及股骨组织形态的影响[J]. 陕西中医, 2025, 46(9): 1172-1177. |
| [19] |
鲁林, 方虹. 骨碎补总黄酮抑制Notch1/Hes1/Prdx1通路对骨质疏松大鼠抗氧化能力的影响[J]. 中国骨质疏松杂志, 2023, 29(10): 1431-1436. |
| [20] |
ZENG L, YANG T, YANG K, et al. Efficacy and safety of curcumin and curcuma longa extract in the treatment of arthritis: A systematic review and Meta-Analysis of randomized controlled trial[J]. Frontiers in Immunology, 2022, 13(1): 891822-891841. |
| [21] |
JIN Z, CHANG B, WEI Y, et al. Curcumin exerts chondroprotective effects against osteoarthritis by promoting AMPK/PINK1/Parkin-mediated mitophagy[J]. Biomedicine & Pharmacotherapy, 2022, 151(1): 113092-113102. |
| [22] |
WANG X, YU H, ZHANG Y, et al. Curcumin alleviates osteoarthritis through the p38MAPK pathway: Network pharmacological prediction and experimental confirmation[J]. Journal of Inflammation Research, 2024, 17(1): 5039-5056. |
| [23] |
CHEN T, ZHOU R, CHEN Y, et al. Curcumin ameliorates IL-1β-induced apoptosis by activating autophagy and inhibiting the NF-κB signaling pathway in rat primary articular chondrocytes[J]. Cell Biology International, 2021, 45(5): 976-988. DOI:10.1002/cbin.11541 |
| [24] |
LI J, LIU W, WANG T, et al. The mechanism of curcumin protecting against IL-1β-induced oxidative stress and inflammation in chondrocytes via the Bmp2/Smad5/Runx2 pathway[J]. Cytotechnology, 2025, 77(2): 71. DOI:10.1007/s10616-025-00731-9 |