 2025, Vol. 44
2025, Vol. 44文章信息
- 郁露露, 袁宇, 刘雪珂, 秦雪彬, 黄丽平, 王玉明, 李遇伯
- YU Lulu, YUAN Yu, LIU Xueke, QIN Xuebin, HUANG Liping, WANG Yuming, LI Yubo
- 20S-原人参三醇改善糖尿病肾病的作用研究
- Effect of 20S-Protopanaxatriol on diabetic kidney disease
- 天津中医药大学学报, 2025, 44(12): 1073-1082
- Journal of Tianjin University of Traditional Chinese Medicine, 2025, 44(12): 1073-1082
- http://dx.doi.org/10.11656/j.issn.1673-9043.2025.12.04
-
文章历史
收稿日期: 2025-08-25

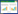



糖尿病肾病(DKD)是常见的糖尿病微血管病变引发的疾病[1],也是全球50%的终末期肾病的致病原因[2]。其发生发展涉及多种相互依赖的危险因素,如肾素-血管紧张素-醛固酮系统和晚期糖基化终产物诱导的炎症、氧化应激及细胞凋亡的激活等[3]。西医治疗DKD主要以对症治疗为主:如降糖、降脂、降压和抗炎疗法[4-5]等。中药长期以来被作为治疗各种重大疾病或慢性疾病的药物来源,在防治复杂疾病方面具有整体调节、副作用小、作用持久等独特优势。已有研究表明,中药中多糖类、多酚类、生物碱类和皂苷类等众多物质可有效改善糖尿病及其并发症[6-7]。例如,黄芪在治疗糖尿病的同时,还被应用于糖尿病心肌损伤、糖尿病血管病变及DKD等并发症的治疗[8];而绞股蓝在治疗DKD的同时,又可改善糖尿病心肌病及糖尿病视网膜病变等[9]。在此背景下,中医为DKD的治疗提供了独特的视角。根据其病因特征,中药或其活性成分以单独或组合的方式从多角度协同作用,产生显著的治疗效果[10-11]。因此,从中药中挖掘DKD的治疗药物具有良好的应用前景。
课题组前期对临床上DKD常用中药进行的中医传承平台分析中,发现DKD治疗以补虚、清热、活血、化瘀为基本治法。补虚药是主要用药类别之一,而人参是其代表药物。人参作为五加科植物人参的干燥根及根茎,有着“百草之王”的美誉,具有大补元气、复脉固脱、健脾益肺、安神益智、生津止渴、补血等众多功效。现已被应用于糖尿病、心血管疾病、肾脏疾病、神经退行性疾病等多种疾病及病症中[12]。在此基础上,结合课题组后期对多种天然产物作用于高糖损伤的人肾皮质近曲小管上皮细胞(HK-2)高通量筛选结果,进一步发现20S-原人参三醇(20S-PPT)对高糖所致的HK-2细胞活力降低具有显著的抑制作用。因此,本研究将建立体内外DKD损伤模型,进一步研究人参中潜在物质基础——20S-PPT改善DKD的作用,为后续相关研究提供数据支撑。
代谢紊乱不仅是DKD的始动因素,更贯穿于DKD进展的整个过程,被认为是DKD发生发展的核心驱动力。近年来,代谢组学作为组学领域的一大分支,能够从生物代谢的直接层面出发,通过反映基因表达变化的功能性后果,有助于更系统地阐明疾病机制或药物作用机制[13]。然而,常规的代谢组学在提供代谢物的空间信息方面缺乏特异性[14],而空间代谢组学作为1种将质谱成像(MSI)与传统代谢组学相结合的新兴技术,弥补了这方面的不足。MSI是1种强大的分子成像技术,可同时对多种代谢物进行原位分析[15]。解吸电喷雾电离(DESI)-MSI是MSI众多技术中的1种,其特点在于能够进行高通量分析,且所需样本量小,可“直接”检测组织切片中的小分子。深入探究差异代谢物的空间特异性改变对DKD进展的影响具有重要的理论和临床意义,故本研究采用DESI-MSI技术,对小鼠肾组织切片进行相关分析,以期发现20S-PPT治疗DKD的生物学机制,也为药物发现提供新的视角。
1 材料 1.1 仪器多功能酶标仪(Thermo Fisher Scientific)、化学发光扫描仪(LI-COR)、流式细胞仪(Beckman Coulter Life Sciences)、摇床(生命科学公司)、CO2恒温孵育箱(Thermo Fisher Scientific)、离心机(Eppendorf)、移液器(大龙医疗设备有限公司)、血糖仪(江苏鱼跃医疗设备股份有限公司)、全自动生化分析仪(日本日立公司)、组织研磨仪(宁波新芝生物科技股份有限公司)、高速离心机(Beckman Coulter Life Sciences)、SYNAPT XS结合DESI-MS质谱成像仪(Waters)、冷冻切片机(徕卡显微系统(上海)贸易有限公司)、涡旋混合器(天悦电子有限公司)。
1.2 主要试剂与药物MEM培养基(武汉普诺赛生命科技有限公司,批号:WHB825N068);CCK-8试剂盒(白鲨生物科技有限公司,批号:23348882);Annexin V-FITC细胞凋亡检测试剂盒(北京索莱宝科技有限公司,批号:23305927);Hoechst 33342活细胞染色液(上海碧云天生物技术有限公司,货号:C1029);20S-PPT标准品(上海源叶生物科技有限公司,含量>99%,批号:P10N9F74763);链脲佐菌霉素(STZ,批号:WXBF0475V);口服葡萄糖(郑州泰丰制药有限公司,国药准字:H41024245,批号:202306024);超氧化物歧化酶(SOD,批号:20240726)、丙二醛(MDA,批号:20240724)、谷胱甘肽过氧化物酶(GSH-Px,批号:20240725)检测试剂盒购自南京建成生物科技有限公司;高脂饲料(小黍有泰生物科技有限公司,货号:D12492)。
1.3 实验动物6周龄雄性C57BL/6J小鼠23只(20~25 g),由北京华阜康实验动物技术有限公司提供,动物许可证号:SYXK(津)2020-0005。饲养于天津中医药大学动物中心,所有动物实验获得本院动物伦理委员会的批准(伦理编号:TCM-LAEC2024085y1518)。于控制温度(25±2)℃、相对湿度为(60%±10%)的环境中饲养,并进行12 h的光照-黑暗循环。
2 方法 2.1 细胞培养及分组常规复苏HK-2细胞,将其培养在MEM(5.5 mmol/L D-葡萄糖)培养基中:90%的MEM培养基、10%的胎牛血清以及1%的双抗,37 ℃,5% CO2。取对数期HK-2细胞,增殖汇合至90%时,胰酶消化,进行传代或冻存。
用含5.5 mmol/L D-葡萄糖的MEM培养基处理细胞,记作对照组(Con组);用含50 mmol/L D-葡萄糖的MEM培养基处理细胞48 h,记作高糖组(HG组);用含50 mmol/L D-葡萄糖的MEM培养基分别配制5、10、25 μmol/L的20S-PPT处理细胞48 h,记作给药组(20S-PPT组)。
2.2 DKD小鼠模型的建立、分组及给药适应性喂养1周后,根据随机数字表法对小鼠进行分组,正常组(Nor组)7只,模型组(Mod组)和给药组(20S-PPT组)各8只。Nor组正常喂养,Mod组及20S-PPT组饲喂高脂饲料,4周后Mod组及20S-PPT组小鼠连续3 d腹腔注射STZ(50 mg/kg),Nor组小鼠注射等体积的柠檬酸钠缓冲液。注射后连续3周使用血糖仪测量小鼠空腹血糖(FBG),FBG>11.1 mmol/L的小鼠被视为糖尿病小鼠。继续高脂喂养4周后,通过代谢笼收集小鼠24 h尿液,检测尿白蛋白和肌酐(CRE),若尿白蛋白/肌酐比值(UACR)与Nor组相比存在显著性差异(P<0.05),则DKD小鼠造模成功[16]。参照文献报道[17-19]的20S-PPT剂量(每天20 mg/kg)给予小鼠,持续灌胃4周。
2.3 检测指标及方法 2.3.1 细胞表型实验细胞活力相关检测:采用CCK-8实验筛选最适给药浓度。随后,取对数期HK-2细胞,增殖汇合至90%时,胰酶消化,MEM培养基重悬至密度为1×105个/mL,接种于6孔板中,各组细胞进行相应处理48 h后,显微镜下观察并用胰酶消化计数。
细胞凋亡相关检测:将HK-2细胞以4×105个/孔接种至6孔板,各组进行相应前处理。为了确定细胞形态变化,根据试剂说明书进行Hoechst染色。用预冷磷酸盐缓冲液(PBS)洗涤细胞后,加入500 μL Hoechst孵育30 min,PBS洗涤细胞3次,在荧光显微镜下进行观察。同等前处理后,收集HK-2并用预冷PBS洗涤,随后按照凋亡试剂盒的说明,用Annexin V-FITC及PI染色后,采用流式细胞仪进行凋亡率的分析。
线粒体功能相关检测:将HK-2接种到6孔板中,对各组细胞进行相应前处理后,根据活性氧(ROS)、线粒体膜电位检测试剂盒的说明,分别进行测定。后续采用Flow Jo及Image J软件进行分析。
2.3.2 小鼠口服糖耐量实验最后1次给药结束后,禁食12 h,对各组小鼠灌胃10%葡萄糖溶液(10 μL/g),在5、15、30、60、120 min检测小鼠血糖值。曲线下面积(AUC)根据以下公式计算:AUC=[0.5×(葡萄糖0+葡萄糖30)+1.5×(葡萄糖30+葡萄糖120)]/2,其中葡萄糖t表示时间t时的葡萄糖浓度。
2.3.3 小鼠体质量增加和器官指数分析检测每日称量各组小鼠体质量,观察体质量变化及小鼠毛发和活动性变化。收集小鼠肾组织样本,记录肾脏湿质量,并计算肾脏系数;测定的器官指数为肾质量(g)/体质量(g)。
2.3.4 小鼠生化分析收集小鼠血浆、尿液及肾组织样本,密封保存至-80 ℃冰箱。收集血样在4 ℃以3 000 r/min离心10 min,离心半径8.4 cm,取上清液检测生化参数,包括尿素氮(BUN)、CRE、尿酸(UA)、总胆固醇(TC)、三酰甘油(TG)、低密度脂蛋白(LDL-C)和高密度脂蛋白(HDL-C);尿液样本用于检测尿白蛋白水平;肾组织样本用于检测SOD、MDA、GSH-Px。
2.3.5 肾脏病理形态学观察将小鼠肾组织样本切成10 μm厚切片,根据标准方案,进行高碘酸-希夫(PAS)染色以观察肾糖原沉积,Masson染色以观察胶原纤维,Tunel染色以检测肾细胞凋亡。DESI-MSI分析后,使用HE染色对组织样本进行组织病理学评估。
2.3.6 小鼠肾组织DESI-MSI分析给药周期结束后,取小鼠肾脏,称质量后放置包埋剂中,-80 ℃保存。使用SYNAPT XS仪器进行分析,该仪器由配备增强DESI源的正交加速(oa)-四极杆(Q)-离子迁移率(IM)-飞行时间(TOF)质谱仪组成,并由MassLynx v4.1控制。使用Leica CM1950紫外线低温恒温器切割10 μm厚的连续切片,并在样品制备前储存在-80 ℃下。溶剂混合物流为乙腈∶水(8∶2,V∶V)。溶剂流量设置为3 μL/min,使用氮气雾化气体喷雾。样品台的运动由Omnispray软件版本2.1.0.2控制。亮氨酸脑啡肽(556.277 1 m/z)用于校准仪器,浓度不超过200 ppb。
为了获得稳定的信号,在每次TOF实验之前,将负离子模式下使用的黑色墨水的响应调整到大约3e6。DESI的几何形状设置如下:入射喷雾角,70 °;喷雾器与入口管居中,-2 mm;喷雾器到入口距离,4 mm;喷雾器到表面的距离,-2 mm。
2.4 统计学方法研究采用SPSS 23.0和GraphPad Prism 9.0软件对数据进行分析及绘图,各组数据以均数±标准差(x±s)表示。对各组数据进行正态分布检验,然后进行方差齐性判断,当各组数据符合正态分布且方差齐时,则采用单因素方差分析(One-Way ANOVA)进行多组间比较,若方差不齐时采用非参数检验,后使用Tukey或Dunnett进行多重比较检验,P<0.05认为结果具有统计学差异。
3 结果 3.1 20S-PPT和高糖对HK-2细胞活力的影响开展表型验证前,进行了20S-PPT的无毒剂量实验,结果说明20S-PPT在浓度40 μmol/L以内,给予细胞48 h无毒性,见图 1A;实验结果显示,20S-PPT在给药浓度5 μmol/L时对高糖损伤HK-2细胞具有显著的保护效果,后续实验选取最低有效剂量进行见图 1 B。

|
| 注:图A,20S-PPT对HK-2细胞的无毒剂量;图B,20S-PPT对HK-2的有效剂量。1,Con组;2,HG组;3,20S-PPT 5 μmol/L组;4,20S-PPT 10 μmol/L组;5,20S-PPT 25 μmol/L组;6,20S-PPT 50 μmol/L组。与Con组相比,*P<0.05,**P<0.01,***P<0.001;与HG组相比,#P<0.05,##P<0.01。 图 1 20S-PPT对HK-2细胞活力的影响(n=3) |
与Con组相比,HG组HK-2细胞形态从鹅卵石状转化为纤维状,细胞数量也显著降低;而20S-PPT组HK-2细胞形态未发生明显改变,且与HG组相比,细胞数量显著增多,见图 2 A-B。Hoechst染色和Annexin V/PI染色结果显示,与Con组相比,HG组HK-2细胞凋亡率显著升高;与HG组相比,20S-PPT组的HK-2细胞凋亡率从11.3%降低至7.3%,提示20S-PPT可抑制HK-2细胞的凋亡,见图 2C-D。流式细胞术及DCFH-DA染色部分结果显示,与Con组相比,高糖损伤的HK-2细胞中ROS水平升高;与HG组相比,20S-PPT组细胞ROS水平显著降低,见图 2 E;荧光强度分析结果同上,见图 2F。最后,对各组细胞进行线粒体膜电位相关检测。结果显示,与Con组相比,HG组HK-2细胞中线粒体膜电位下降;与HG组相比,20S-PPT给药组细胞中线粒体膜电位水平得到回调, 见图 2G。

|
| 注:图A,各组细胞形态变化;图B,各组细胞数量变化;图C,各组细胞Hoechst染色情况;图D,流式细胞术检测各组细胞凋亡情况;图E,流式细胞术检测各组细胞内ROS变化;图F,各组细胞DCFH-DA染色情况;图G,各组细胞线粒体膜电位变化情况。与Con组相比,*P<0.05,**P<0.01;与HG组相比,#P<0.05;##P<0.01。 图 2 20S-PPT对高糖损伤HK-2细胞的保护作用(n=3) |
在本研究中,造模成功率为100%,Nor组存活率为100%,Mod组存活率为87.5%。与Nor组相比,Mod组小鼠的体质量减少,饮水量、FBG、口服糖耐量(OGTT)和肾指数明显升高;20S-PPT治疗4周后,与Mod组相比,20S-PPT组小鼠体质量显著升高,饮水量、FBG、OGTT、肾指数显著降低,见图 3A-E。此外,与Nor组相比,Mod组小鼠的24 hUACR水平显著升高,见图 3F,揭示了肾损伤形成。如图 3 G所示,Nor组小鼠肾组织结构完整,未发生异常病理学改变;Mod组小鼠肾组织整体结构异常,可见肾小球及肾小管,间质中度扩张,褐色凋亡小点,肾小球囊及系膜可见糖原沉积堆积;经20S-PPT干预后,小鼠肾组织中的糖原沉积、肾细胞凋亡、胶原蛋白积累和纤维化情况得到改善。此外,与Mod组相比,20S-PPT治疗后小鼠CRE、BUN、UA、TC、TG、LDL-C及MDA水平明显下降,HDL-C及SOD和GSH-PX水平明显上升,见图 3H-Q。提示20S-PPT治疗显著改善小鼠糖脂失调和氧化应激水平。

|
| 注:图A,体质量变化;图B,饮水量变化;图C,FBG变化;图D,OGTT变化及曲线下面积;图E,肾指数水平;图F,UACR水平;图G,肾脏病理变化;图H,CRE水平;图I,BUN水平;图J,UA水平;图K,TC水平;图L,TG水平;图M,LDL-C水平;图N,HDL-C水平;图O,SOD水平;图P,MDA水平;图Q,GSH-PX水平。与Nor组相比,*P<0.05,**P<0.01,***P<0.001;与Mod组相比,#P<0.05,##P<0.01,###P<0.001。 图 3 20S-PPT对DKD小鼠的肾脏保护作用(n=6) |
对各组小鼠肾脏切片进行DESI-MSI分析。结果显示,治疗后共34种注释的内源性代谢物的丰度和组织分布发生了明显变化(ppm<5),22种代谢物的丰度呈增加趋势(4种代谢物显著增加,P<0.05),11种代谢物的丰度呈降低趋势(5种代谢物显著降低,P<0.05),见表 1。这表明20S-PPT治疗后,肾组织的代谢发生了高度变化,主要涉及葡萄糖代谢、三羧酸循环(TCA)、核苷酸代谢及脂质代谢等,具体内容见开放科学(资源服务)标识码(OSID)。
| 代谢物英文名称 | 代谢物中文名称 | 结构式 | 加合方式 | 理论值 m/z |
测量值 m/z |
ppm | MS/MS | DKD(vs Nor组) | 20S-PPT(vs Mod组) |
| Malic acid | 苹果酸 | C4H6O5 | [M-H]- | 133.013 7 | 133.013 0 | 5.26 | 115,133 | ↓* | ↑# |
| Hypoxanthine | 次黄嘌呤 | C4H8O5 | [M-H]- | 135.029 3 | 135.029 6 | 2.22 | 65,92 | ↓ | ↑# |
| DL-Glutamate | 谷氨酸钠 | C5H9NO4 | [M-H]- | 147.053 2 | 147.045 0 | 5.57 | 102,130,148 | ↓** | ↑ |
| Xanthine | 黄嘌呤 | C5H4N4O2 | [M-H]- | 151.025 6 | 151.025 0 | 3.97 | 80,108 | ↓* | ↑# |
| Succinic acid | 琥珀酸 | C4H6O4 | [M+Cl]- | 152.995 5 | 152.994 6 | 5.88 | 73,99,117 | ↑* | ↓# |
| Uric acid | 尿酸 | C5H4N4O3 | [M-H]- | 167.020 5 | 167.019 9 | 3.59 | 69,96 | ↓ | ↑ |
| Ascorbic acid | 维生素C | C6H8O6 | [M-H]- | 175.024 3 | 175.023 7 | 3.40 | 87 | ↓ | ↑ |
| Inosine | 肌苷 | C10H12N4O5 | [M-H]- | 267.072 9 | 267.072 3 | 2.24 | 137 | ↓ | ↑ |
| Glycerylphosphorylethanolamine | 甘油磷酰乙醇胺 | C5H14NO6P | [M-H]- | 214.048 0 | 214.047 2 | 3.73 | 214 | ↓ | ↑ |
| D-Glucose | D-葡萄糖 | C6H12O6 | [M+Cl]- | 215.032 2 | 215.031 8 | 1.86 | 59,71,89 | ↑ | ↓# |
| Citric acid | 柠檬酸 | C6H8O7 | [M-H]- | 191.019 2 | 191.018 1 | 5.75 | 85,87,110 | ↓* | ↑ |
| Uridine | 尿苷 | C9H12N2O6 | [M-H]- | 243.061 7 | 243.061 8 | 4.11 | 110,243 | ↓* | ↑ |
| Fructose 6-phosphate | 果糖6-磷酸 | C6H13O9P | [M-H]- | 259.021 9 | 259.021 1 | 3.08 | 259 | ↑ | ↓ |
| Linoleic acid | 亚油酸 | C18H32O2 | [M-H]- | 279.232 4 | 279.231 7 | 2.50 | 279 | ↓* | ↑ |
| Oleic acid | 油酸 | C18H34O2 | [M-H]- | 281.248 1 | 281.247 0 | 3.91 | 88,196,281 | ↑ | ↓# |
| Arachidonic acid | 花生四烯酸 | C20H32O2 | [M-H]- | 303.232 4 | 303.231 7 | 2.30 | 259,303 | ↓ | ↑ |
| Docosahexaenoic acid | 二十二碳六烯酸 | C22H32O2 | [M-H]- | 327.232 4 | 327.231 6 | 2.44 | 67,238,309,327 | ↓* | ↑ |
| Guanosine monophosphate(GMP) | 鸟苷酸 | C10H14N5O8P | [M-H]- | 362.050 2 | 362.048 8 | 3.86 | 79,97,150,362 | ↑* | ↓ |
| Cytidine | 胞苷 | C9H13N3O5 | [M-H]- | 242.077 7 | 242.078 5 | 3.30 | 112,243 | ↓ | ↑ |
| Adenosine diphosphate(ADP) | 二磷酸腺苷 | C10H15N5O10P2 | [M-H]- | 426.021 6 | 426.020 3 | 3.05 | 78,328,426 | ↑* | ↓ |
| PE(P-38:6) | - | C43H74NO7P | [M-H]- | 746.512 5 | 746.510 3 | 2.94 | 196,327,436,746 | ↓ | ↑ |
| PE(38:6) | - | C43H74NO8P | [M-H]- | 762.507 4 | 762.506 1 | 1.70 | 256,327,484 | ↓ | ↑ |
| PE(38:4) | - | C43H78NO8P | [M-H]- | 766.538 7 | 766.536 9 | 2.34 | 766 | ↓ | ↑ |
| Glycerophosphoric acid | 甘油磷酸 | C3H9O6P | [M-H]- | 171.005 8 | 171.006 0 | 1.16 | 79,97,171,214 | ↓ | ↑ |
| LysoPE(18:0/0:0) | - | C23H48NO7P | [M-H]- | 480.309 0 | 480.308 0 | 2.08 | 196,480 | ↓** | ↑ |
| PC(18:0/20:4) | - | C46H82NO9P | [M-H]- | 822.564 9 | 822.567 3 | 2.91 | 184,209 | ↓ | ↑# |
| D/L-Histidine | D-组氨酸 | C6H9N3O2 | [M-H]- | 154.061 7 | 154.061 2 | 3.24 | 93,110 | ↑ | ↓ |
| Glutathione | 谷胱甘肽 | C10H17N3O6S | [M-H]- | 306.076 0 | 306.074 6 | 4.57 | 80,123,272 | ↑* | ↓ |
| PC(35:4) | - | C43H78NO8P | [M-H]- | 767.546 5 | 767.540 2 | 6.25 | 211 | ↑* | ↓# |
| PS(38:6) | - | C44H74NO10P | [M-H]- | 808.512 9 | 808.512 0 | 1.11 | 255,327 | ↑** | ↓## |
| PGP(20:4) | - | C46H78O17P2 | [M-H]- | 963.463 6 | 963.466 3 | 2.80 | - | ↓* | ↑ |
| PGP(20:3) | - | C46H80O17P2 | [M-H]- | 966.487 1 | 966.485 1 | 2.06 | - | ↓ | ↑ |
| PA(25:0) | - | C41H73O8P | [M-H]- | 723.496 5 | 723.494 7 | 2.48 | - | ↑** | ↓# |
| Adenosine monophosphate(AMP) | 腺苷酸单磷酸盐 | C10H14N5O7P | [M-H]- | 346.055 3 | 346.054 1 | 3.46 | 78,97,134,211,346 | ↑ | ↓ |
| 注:与Nor组相比,*P<0.05,**P<0.01;与Mod组相比,#P<0.05,##P<0.01。↑:呈增加趋势,↓:呈降低趋势。 | |||||||||
人参皂苷是人参的主要活性成分,可通过调节胰岛素分泌、调节脂质代谢、抗糖、抗氧化应激和抗炎发挥治疗糖尿病及并发症的作用[20],其活性成分主要包括人参皂苷Rg1、Rg3、Rb1、Rb3、Rh2、Rc、Re等[21]。研究表明,口服摄入的人参皂苷Rg1通过肠道得到Rh1和F1,继而被结肠细菌去糖基化,最终获得主要代谢物20S-原人参二醇20-O-β-D-吡喃葡萄糖苷和20S-PPT[22]。经证实,与人参皂苷Rg1相比,其次级代谢产物20S-PPT具有更强的生物活性。现代药理学证明,20S-PPT具有抗炎、抗氧化、抗脂肪、改善胰岛素抵抗的作用[23],其具体的作用机制包括:1)特异性抑制炎症介导的炎症小体活化。2)直接或间接抑制NF-κB的活性,诱导炎症介质IL-1β、诱导型一氧化氮合酶、环加氧酶2和一氧化氮的生成[24]。3)可剂量依赖性地增加过氧化物酶体增殖物激活受体反式活化,从而改善与糖尿病相关的胰岛素抵抗[25]。4)影响葡萄糖转运体4表达,改善葡萄糖稳态[25]。此外,还可起到改善血脂代谢、抑制系膜细胞的增殖和分化[22]等作用。因此,本研究拟通过构建DKD体内外模型,初步探讨20S-PPT对该疾病的影响。
4.2 核苷酸代谢紊乱与DKD的相关性代谢组学是识别新的预后生物标志物、治疗靶点和中药在疾病治疗中分子机制的有力策略[26]。而空间代谢组学可从三维角度分析不同组织与器官中的代谢物变化,为中药代谢物组织特异性定位提供有价值的见解。研究借助DESI-MSI技术发现,20S-PPT治疗后共34种注释的内源性代谢物的丰度和组织分布发生了显著变化,并涉及多条代谢通路,包括核苷酸代谢、葡萄糖代谢、TCA循环、脂质代谢等。其中,核苷酸代谢包括嘌呤代谢及嘧啶代谢,该途径已被证实与DKD的进展息息相关。在本研究中,20S-PPT治疗DKD过程中涉及的核苷酸类代谢物主要有尿苷、黄嘌呤、肌苷、胞苷等。对嘧啶类化合物的代谢和作用的研究已经揭示了大量关于这些内源性代谢物如何调节细胞生理学的信息。嘧啶作为所有活细胞的重要组成部分,它们具有广泛的多种生物功能,尤其是调节细胞代谢、能量守恒和运输、辅酶和磷脂活性中间体的形成以及碳水化合物代谢[27]。嘧啶代谢紊乱可通过影响磷脂的合成代谢或直接损伤肾内血管的内皮细胞,导致肾血流动力学异常,从而驱动糖尿病和DKD的发生[28-29]。而嘌呤代谢的紊乱常表现在血、尿中尿酸水平的变化。尿酸作为嘌呤代谢的终端产物,具有强大的抗氧化作用,通常依赖于肾脏排泄,在诱导肾脏炎症、肾内血管收缩和肾损伤中起重要作用,可导致急性肾功能衰竭或高尿酸血症[30-31]。
4.3 糖代谢紊乱与DKD的相关性在DKD损伤过程中,肾脏发生糖代谢重编程,促进氧化和纤维化。在体内,糖代谢涉及糖的无氧氧化和有氧氧化,包含糖酵解、TCA循环、丙酮酸代谢和单糖转化。在本研究中,DKD主要涉及的相关代谢物有果糖-6-磷酸、苹果酸和柠檬酸等。TCA循环是所有生物体中存在的基本代谢途径,也是主要发生在线粒体中的能量代谢关键途径[32],该循环依赖于柠檬酸盐、富马酸盐和苹果酸盐等中间代谢物[33]。Li等[34]研究者对db/db小鼠的TCA循环代谢扰动进行了长期的监测。发现在DKD早期,小鼠血清中柠檬酸盐、苹果酸盐的血清水平显著升高,说明TCA循环周期大大加速,表明此阶段葡萄糖代谢正常。而在DKD后期,小鼠体内葡萄糖水平逐渐升高,柠檬酸盐与苹果酸盐的水平显著降低,表明进入TCA循环的葡萄糖较少或通过TCA循环代谢葡萄糖的效率较低,从而进一步增加血糖水平。而琥珀酸盐是唯一表现出与其他中间体不同趋势的TCA中间体,这与本研究结果相符。
4.4 脂质代谢紊乱与DKD的相关性除此之外,由脂质积累及其代谢物驱动的脂毒性也是DKD发病机制的关键因素之一[35]。当DKD患者出现肾损伤时,随着尿白蛋白排泄的增加,被肾小球过滤的脂肪酸也增加,导致与白蛋白结合的过量游离脂肪酸超负荷并在肾小管中被重吸收,这将导致严重的肾小管间质损伤[36]。值得一提的是,亚油酸和亚麻酸是具有抗炎特性的必需多不饱和脂肪酸。适当摄入这些多不饱和脂肪酸对改善DKD患者的肾功能起着积极作用[37]。而本研究结果显示,20S-PPT干预后小鼠肾组织中亚油酸和亚麻酸水平明显升高,与上述报道相符。此外,磷脂是包含多个脂肪酰基的所有细胞脂质双层的关键结构成分,已被认定为DKD的潜在生物标志物[38]。磷脂酰丝氨酸(PS)、磷脂酰胆碱(PC)和磷脂酸(PA)在病理状态下发生显著改变[39],其异常代谢通过影响细胞膜完整性、信号传导、炎症反应和纤维化等过程,直接参与肾脏损伤的多个环节。多项研究发现脂质积累推动DKD的发展(蛋白尿和糖尿病肾小球硬化)与损伤肾小球机制、引发代谢重编程效应等相关,导致脂质的紊乱与DKD形成“代谢-炎症-纤维化”的恶性循环。20S-PPT可以调节上述代谢紊乱,这可能为20S-PPT防治DKD提供新的治疗策略。
4.5 不足与局限性最后,本研究还存在些许不足。首先,动物数量较少且未设置阳性药物组,无法评估20S-PPT与现有疗法的优劣性,只能够初步明确20S-PPT对DKD的治疗效果;其次,DKD存在中医体质的易感性,不同的体质会导致DKD多样性证候,明确体质特征,有助于更好的阐明药物作用于疾病的治疗原理[40-41]。再次,20S-PPT作为补虚药人参的代谢产物,应基于病证结合深入挖掘其治疗DKD具体证型的作用及机制。其中,“病”与“证”的同步性、模型的稳定性与重复性、证候的特异性等问题都需要仔细斟酌[42]。
5 结论研究通过体外与体内实验系统阐明了20S-PPT对DKD的保护作用及相关分子机制。体外实验表明,20S-PPT在5 μmol/L浓度下作用48 h,可显著改善高糖环境诱导的HK-2细胞损伤。具体而言,其通过调控细胞凋亡进程以及缓解氧化应激反应,从而维持HK-2细胞的生物学功能;在动物实验层面,证实20S-PPT以20mg/kg剂量干预DKD模型小鼠后,可有效改善小鼠肾功能及病理损伤;进一步机制研究表明,20S-PPT的上述保护作用与代谢调控密切相关,主要涉及核苷酸代谢中嘌呤与嘧啶的合成调节、糖脂代谢紊乱的改善以及TCA循环中代谢物的积累与转化效率等。整体而言,本研究从细胞、动物和机制多个维度,为20S-PPT后续的临床转化研究及作用机制的深入探讨提供了可靠的实验依据。
| [1] |
Expert consensus on the diagnosis and management of diabetic kidney disease[J]. Chinese Medical Journal, 2020, 133(19): 2333-2334.
|
| [2] |
GUO W C, SONG Y C, SUN Y, et al. Systemic immune-inflammation index is associated with diabetic kidney disease in Type 2 diabetes mellitus patients: Evidence from NHANES 2011-2018[J]. Frontiers in Endocrinology, 2022, 13: 1071465. DOI:10.3389/fendo.2022.1071465 |
| [3] |
AKHTAR M, TAHA N M, NAUMAN A, et al. Diabetic kidney disease: Past and present[J]. Advances in Anatomic Pathology, 2020, 27(2): 87-97. DOI:10.1097/PAP.0000000000000257 |
| [4] |
ZHAO J, TOSTIVINT I, XU L D, et al. Efficacy of combined Abelmoschus manihot and irbesartan for reduction of albuminuria in patients with type 2 diabetes and diabetic kidney disease: A multicenter randomized double-blind parallel controlled clinical trial[J]. Diabetes Care, 2022, 45(7): e113-e115. DOI:10.2337/dc22-0607 |
| [5] |
FINEBERG D, JANDELEIT-DAHM K A M, COOPER M E. Diabetic nephropathy: Diagnosis and treatment[J]. Nature Reviews Endocrinology, 2013, 9(12): 713-723. DOI:10.1038/nrendo.2013.184 |
| [6] |
尚杰, 刘淼, 梁洋, 等. 中药活性成分治疗糖尿病及其并发症的研究进展[J]. 世界科学技术-中医药现代化, 2022, 24(5): 1729-1737. |
| [7] |
王玥彤, 季昭臣, 张雅姿, 等. 中成药治疗糖尿病肾病的随机对照临床试验现状及质量评价[J]. 天津中医药, 2024, 41(9): 1135-1142. |
| [8] |
马艳春, 段莹, 胡建辉, 等. 黄芪治疗糖尿病及其并发症研究进展[J]. 中医药学报, 2022, 50(6): 103-107. |
| [9] |
诸夔妞, 肖洪贺, 寿旗扬, 等. 绞股蓝及其活性成分改善糖尿病并发症作用机制的研究进展[J]. 中国药房, 2020, 31(8): 1015-1020. |
| [10] |
LI H Y, ZHAO X L, ZHENG L Y, et al. Bruceine A protects against diabetic kidney disease via inhibiting galectin-1[J]. Kidney International, 2022, 102(3): 521-535. DOI:10.1016/j.kint.2022.04.020 |
| [11] |
DONG R X, ZHANG X, LIU Y D, et al. Rutin alleviates EndMT by restoring autophagy through inhibiting HDAC1 via PI3K/AKT/mTOR pathway in diabetic kidney disease[J]. Phytomedicine, 2023, 112: 154700. DOI:10.1016/j.phymed.2023.154700 |
| [12] |
AHMAD S S, AHMAD K, HWANG Y C, et al. Therapeutic applications of ginseng natural compounds for health management[J]. International Journal of Molecular Sciences, 2023, 24(24): 17290. DOI:10.3390/ijms242417290 |
| [13] |
TENNANT D A, DURáN R V, GOTTLIEB E. Targeting metabolic transformation for cancer therapy[J]. Nature Reviews Cancer, 2010, 10(4): 267-277. DOI:10.1038/nrc2817 |
| [14] |
BAN W K, YOU Y Y, YANG Z H. Imaging technologies for cerebral pharmacokinetic studies: Progress and perspectives[J]. Biomedicines, 2022, 10(10): 2447. DOI:10.3390/biomedicines10102447 |
| [15] |
SPENGLER B. Mass spectrometry imaging of biomolecular information[J]. Analytical Chemistry, 2015, 87(1): 64-82. DOI:10.1021/ac504543v |
| [16] |
刘懿娴, 李佳琪, 黄鹭铭, 等. 积雪草酸抑制TXNIP/NLRP3信号通路改善小鼠糖尿病肾病的机制研究[J]. 中药新药与临床药理, 2022, 33(12): 1614-1622. |
| [17] |
SU W Y, LI Y, CHEN X, et al. Ginsenoside Rh1 improves type 2 diabetic nephropathy through AMPK/PI3K/Akt-mediated inflammation and apoptosis signaling pathway[J]. The American Journal of Chinese Medicine, 2021, 49(5): 1215-1233. DOI:10.1142/S0192415X21500580 |
| [18] |
FANG S, CAI Y Y, LYU F P, et al. Exendin-4 improves diabetic kidney disease in C57BL/6 mice independent of brown adipose tissue activation[J]. Journal of Diabetes Research, 2020, 2020(1): 9084567. |
| [19] |
WANG C Z, TAN L Y, LIU J T, et al. Integrated metabolomics and network pharmacology to decipher the latent mechanisms of protopanaxatriol against acetic acid-induced gastric ulcer[J]. International Journal of Molecular Sciences, 2022, 23(20): 12097. DOI:10.3390/ijms232012097 |
| [20] |
高健, 吕邵娃. 人参化学成分及药理作用研究进展[J]. 中医药导报, 2021, 27(1): 127-130, 137. |
| [21] |
王丽杰, 王春艳, 张冬燕, 等. 人参治疗糖尿病及作用机制研究进展[J]. 中国热带医学, 2013, 13(7): 906-909. |
| [22] |
HASEGAWA H. Proof of the mysterious efficacy of ginseng: Basic and clinical trials: Metabolic activation of ginsenoside: Deglycosylation by intestinal bacteria and esterification with fatty acid[J]. Journal of Pharmacological Sciences, 2004, 95(2): 153-157. DOI:10.1254/jphs.FMJ04001X4 |
| [23] |
陈小玲, 史大臻, 司函瑞, 等. 原人参三醇生物活性及其作用机制研究进展[J]. 辽宁中医药大学学报, 2021, 23(2): 209-214. |
| [24] |
JIN L N, FU X, YAO S S, et al. Protective effects of protopanaxatriol on acute liver injury induced by concanavalin A[J]. Naunyn-Schmiedeberg's Archives of Pharmacology, 2019, 392(1): 81-87. DOI:10.1007/s00210-018-1567-4 |
| [25] |
HAN K L, JUNG M H, SOHN J H, et al. Ginsenoside 20S-protopanaxatriol(PPT) activates peroxisome proliferator-activated receptor gamma(PPARgamma) in 3T3-L1 adipocytes[J]. Biological & Pharmaceutical Bulletin, 2006, 29(1): 110-113. |
| [26] |
HAN J W, LI P, SUN H, et al. Integrated metabolomics and mass spectrometry imaging analysis reveal the efficacy and mechanism of Huangkui capsule on type 2 diabetic nephropathy[J]. Phytomedicine, 2025, 138: 156397. DOI:10.1016/j.phymed.2025.156397 |
| [27] |
XIA J F, HU P, LIANG Q L, et al. Correlations of creatine and six related pyrimidine metabolites and diabetic nephropathy in Chinese type 2 diabetic patients[J]. Clinical Biochemistry, 2010, 43(12): 957-962. DOI:10.1016/j.clinbiochem.2010.05.013 |
| [28] |
LIU C X, GU C Y, HUANG W, et al. Targeted UPLC-MS/MS high-throughput metabolomics approach to assess the purine and pyrimidine metabolism[J]. Journal of Chromatography B, Analytical Technologies in the Biomedical and Life Sciences, 2019, 1113: 98-106. DOI:10.1016/j.jchromb.2019.03.008 |
| [29] |
HOLGATE S T, WENZEL S, POSTMA D S, et al. Asthma[J]. Nature Reviews Disease Primers, 2015, 1: 15025. DOI:10.1038/nrdp.2015.25 |
| [30] |
ROMI M M, ARFIAN N, TRANGGONO U, et al. Uric acid causes kidney injury through inducing fibroblast expansion, Endothelin-1 expression, and inflammation[J]. BMC Nephrology, 2017, 18(1): 326. DOI:10.1186/s12882-017-0736-x |
| [31] |
STEELE T H. Hyperuricemic nephropathies[J]. Nephron, 1998, 81(Suppl. 1): 45-49. |
| [32] |
TAN S M, ZIEMANN M, THALLAS-BONKE V, et al. Complement C5a induces renal injury in diabetic kidney disease by disrupting mitochondrial metabolic agility[J]. Diabetes, 2020, 69(1): 83-98. DOI:10.2337/db19-0043 |
| [33] |
MARTÍNEZ-REYES I, CHANDEL N S. Mitochondrial TCA cycle metabolites control physiology and disease[J]. Nature Communications, 2020, 11: 102. DOI:10.1038/s41467-019-13668-3 |
| [34] |
LI M J, WANG X F, AA J, et al. GC/TOFMS analysis of metabolites in serum and urine reveals metabolic perturbation of TCA cycle in db/db mice involved in diabetic nephropathy[J]. American Journal of Physiology Renal Physiology, 2013, 304(11): F1317-F1324. DOI:10.1152/ajprenal.00536.2012 |
| [35] |
THONGNAK L, PONGCHAIDECHA A, LUNGKAPHIN A. Renal lipid metabolism and lipotoxicity in diabetes[J]. The American Journal of the Medical Sciences, 2020, 359(2): 84-99. DOI:10.1016/j.amjms.2019.11.004 |
| [36] |
KAMIJO A, KIMURA K, SUGAYA T, et al. Urinary free fatty acids bound to albumin aggravate tubulointerstitial damage[J]. Kidney International, 2002, 62(5): 1628-1637. DOI:10.1046/j.1523-1755.2002.00618.x |
| [37] |
DOS SANTOS A L T, DUARTE C K, SANTOS M, et al. Low linolenic and linoleic acid consumption are associated with chronic kidney disease in patients with type 2 diabetes[J]. PLoS One, 2018, 13(8): e0195249. DOI:10.1371/journal.pone.0195249 |
| [38] |
ZHAO Y Y, VAZIRI N D, LIN R C. Lipidomics: New insight into kidney disease[J]. Advances in Clinical Chemistry, 2015, 68: 153-175. |
| [39] |
KARPE F, DICKMANN J R, FRAYN K N. Fatty acids, obesity, and insulin resistance: Time for a reevaluation[J]. Diabetes, 2011, 60(10): 2441-2449. DOI:10.2337/db11-0425 |
| [40] |
刘传鑫, 孔娇. 体质毒理学: 中药安全性评价的新方向[J]. 世界科学技术-中医药现代化, 2023, 25(12): 3776-3784. |
| [41] |
张宁, 赵泉霖. 基于"体质-疾病-证候"模式的糖尿病肾病防治思路[J]. 中医杂志, 2018, 59(22): 1920-1923. |
| [42] |
许艳超, 郑曙琴. 基于数据挖掘技术的糖尿病肾病微量蛋白尿期与临床蛋白尿期证候比较研究[J]. J天津中医药大学学报, 2024, 43(11): 1017-1020. |