 2025, Vol. 44
2025, Vol. 44文章信息
- 王易, 张亚美, 向蓓蓓
- WANG Yi, ZHANG Yamei, XIANG Beibei
- 硝普钠对藏药川西獐牙菜生长及药用成分积累的影响
- Effect of sodium nitroprusside on the growth and accumulation of medicinal components of Swertia mussotii Franch
- 天津中医药大学学报, 2025, 44(4): 309-316
- Journal of Tianjin University of Traditional Chinese Medicine, 2025, 44(4): 309-316
- http://dx.doi.org/10.11656/j.issn.1673-9043.2025.04.05
-
文章历史
收稿日期: 2025-01-13




川西獐牙菜(Swertia mussotii Franch.)是隶属龙胆科(Gentianaceae)獐牙菜属(Swertia)的一年生草本植物,也是藏茵陈的基原植物之一[1]。川西獐牙菜以干燥全草入药,具有巨大的经济和药用价值。《晶珠本草》记载川西獐牙菜具有清肝、利尿、通筋骨、止血等功效,目前在临床上常用于治疗肝胆疾病[2]。根据相关研究报道,獐牙菜苦苷、芒果苷、龙胆苦苷、獐牙菜苷和齐墩果酸是川西獐牙菜的主要活性成分[3]。这类呫吨酮及环烯醚萜类成分具有抗肿瘤、抗炎利胆、降血糖、护肝、抗病毒等功效[4]。然而川西獐牙菜药材资源稀少且有效成分提取困难,因此增加川西獐牙菜中有效成分的含量对于提高其药用价值具有重要意义。
一氧化氮(NO)是一种小型气体生物活性信号分子,参与多种生物过程[5]。NO是启动植物次生代谢产物生物合成的信号转导途径中的关键分子,能够诱导植物次生代谢产物的积累。硝普钠(SNP)溶解后能够释放NO,是一种常用的NO供体[6]。外源施加SNP可以延缓植物衰老,同时激活或抑制植物次生代谢途径相关基因的表达,进一步影响药用成分的合成[7]。研究表明,SNP能够诱导植物中多酚[8]、灵芝三萜[9]、青蒿素[10]和紫杉醇[11]的生物合成。但是,目前尚未有SNP处理后川西獐牙菜生长及药用成分的相关报道。本研究探讨外源施加不同浓度SNP对川西獐牙菜生长、光合色素含量、抗氧化以及次生代谢产物积累的影响,为将来应用外源物质促进川西獐牙菜生长及调控药用成分积累提供思路。
1 材料与方法 1.1 实验材料 1.1.1 供试材料川西獐牙菜种子采集于青海省玉树藏族自治州,经青海大学董汇泽教授鉴定为川西獐牙菜种子。
1.1.2 供试药剂SNP(solarbio);核黄素(solarbio);三氯乙酸(solarbio);氯化硝基氮蓝四唑(solarbio);无水乙醇(国药集团化学试剂有限公司);硫代巴比妥酸(国药集团化学试剂有限公司);乙二胺四乙酸二钠(EDTA-2Na,上海阿拉丁生化科技股份有限公司);蛋氨酸(上海源叶生物科技有限公司);愈创木酚(上海源叶生物科技有限公司);过氧化氢(山东利尔康医疗科技股份有限公司);磷酸氢二钠(天津市华东试剂厂);磷酸二氢钠(天津市光复科技发展有限公司);甲醇(色谱纯,Sigma);乙腈(色谱纯,Sigma);甲酸(上海麦克林生化科技有限公司);液相用水(屈臣氏);獐牙菜苦苷标准品(成都瑞芬思生物科技有限公司,批号:RFS-Z00211808004);獐牙菜苷标准品(成都瑞芬思生物科技有限公司,批号:RFS-Z00902112017);龙胆苦苷标准品(成都瑞芬思生物科技有限公司,批号:RFS-L00502112015);芒果苷标准品(成都瑞芬思生物科技有限公司,批号:RFS-M01802011021);异荭草素标准品(成都瑞芬思生物科技有限公司,批号:RFS-Y13211805031)。
1.1.3 供试仪器超高效液相色谱仪ACQUITY UPLC BEH C18型(美国waters公司);超声波清洗机KS-500DE型(昆山洁力美超声仪器有限公司);电热恒温鼓风干燥箱DHG-9240B型(上海培因实验仪器有限公司);高速冷冻离心机ST 16R型(赛默飞世尔科技有限公司);水浴锅HH-S2型(常州润华电器有限公司);紫外可见分光光度计Alpha-1860A型(上海谱元仪器有限公司);-80 ℃冷藏箱TSU500V型(赛默飞世尔科技有限公司);荧光定量PCR仪Bio-Rad型[伯乐生命医学产品(上海)有限公司];十万分之一天平ME215S型(德国Satorius公司);叶绿素荧光仪RS232型(汉莎科技集团有限公司)。
1.2 实验方法川西獐牙菜种子在双蒸水中浸泡12 h后播种于无菌土中,在温度(25±1)℃、光照16 h+黑暗8 h循环的条件下培养获得3月龄的土培苗。精密称取SNP粉末,用蒸馏水配制成最终浓度为0.5、1.0、2.0、3.0 mmol/L的SNP溶液,对长势相近的3月龄土培苗隔日进行喷洒处理,同时设置等量蒸馏水喷洒处理为对照组(即0 mmol/L)。每个浓度平行处理10株土培苗,共计处理10 d。10 d后收样,进行后续指标测定。
1.3 项目测定 1.3.1 鲜、干质量测定用蒸馏水冲洗土培苗去除杂质后擦干并称取质量,即为川西獐牙菜的鲜质量。80 ℃恒温烘干至质量恒定并称取质量,即为川西獐牙菜的干质量。每个浓度平行测定3株土培苗。
1.3.2 光合色素含量测定称取50.0 mg新鲜土培苗于5 mL EP管中进行光合色素含量测定,测量方法参考Zhang等[12]的描述。每个浓度样品平行测定3次。
1.3.3 叶绿素荧光参数测定通过叶绿素荧光诱导动力学曲线能够快速了解植物体内光系统Ⅱ(PSⅡ)的原初光化学反应及光合机构中电子传递状态的变化。选取不同浓度SNP处理后长势一致、状态良好的川西獐牙菜叶片暗处理30 min后进行测量,每个浓度平行测量3株土培苗。
1.3.4 丙二醛(MDA)含量及抗氧化酶活性测定称取0.1 g新鲜的土培苗于2 mL EP管中进行MDA含量和超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性测定,测量方法参考Zhang等[12]的描述。每个浓度样品平行测定3次。
1.3.5 川西獐牙菜药效成分含量测定 1.3.5.1 色谱条件色谱柱:ACQUITY UPLC BEH C18(2.1 mm×50 mm,1.7 μm);检测波长:250 nm;流速:0.3 mL/min;柱温:30 ℃;上样体积:1 μL;流动相:0.1%甲酸水溶液(A)-纯乙腈(B)。梯度洗脱(0~7 min,10%~30%B;7~8 min,30%~10%B;8~10 min,10%B)。色谱图见图 1。

|
| 注:图A,混合对照品;图B,供试品。1,獐牙菜苦苷;2,芒果苷;3,龙胆苦苷;4,獐牙菜苷;5,异荭草素。 图 1 川西獐牙菜的超高效液相色谱(UPLC)图 |
称取川西獐牙菜干燥粉末50.0 mg,加入甲醇10 mL,50 ℃超声提取50 min。放冷后补足缺失质量,4 ℃,12 000 r/min,离心半径250 mm,离心10 min。经0.22 μm微孔滤膜过滤,取续滤液作为供试品溶液,4 ℃保存备用。
1.3.5.3 对照品溶液制备分别精密称定獐牙菜苦苷、芒果苷、龙胆苦苷、獐牙菜苷和异荭草素标准品1.52、2.20、6.05、1.51、1.13 mg于5 mL容量瓶中,加入甲醇超声溶解,定容至5 mL,制成浓度分别为304、440、1 210、302、226 μg/mL的单一对照品储备液,4 ℃保存备用。分别精密吸取上述单一对照品溶液各1 mL于5 mL容量瓶中,定容至5 mL,得到獐牙菜苦苷60.8 μg/mL、芒果苷88.0 μg/mL、龙胆苦苷242.0 μg/mL、獐牙菜苷60.4 μg/mL和异荭草素45.2 μg/mL的混合对照品溶液。
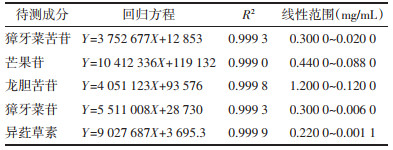
1.3.5.4 标准曲线绘制分别精密吸取獐牙菜苦苷、芒果苷、龙胆苦苷、獐牙菜苷和异荭草素各对照品溶液,加入甲醇逐级稀释后进样测定。以质量浓度为横坐标,色谱峰面积为纵坐标,绘制线性回归曲线。结果显示獐牙菜苦苷、芒果苷、龙胆苦苷、獐牙菜苷、异荭草素的线性关系良好。见表 1。
精密吸取混合对照品溶液,连续进样6次,记录峰面积。结果显示獐牙菜苦苷、芒果苷、龙胆苦苷、獐牙菜苷、异荭草素峰面积的RSD值分别为1.92%、1.07%、1.28%、1.98%、1.58%,说明UPLC色谱仪精密度良好。
1.3.5.6 稳定性实验吸取同一供试品溶液,在0、2、4、6、8、10、12 h进样,记录峰面积,12 h内各成分峰面积的RSD值分别为1.43%、1.28%、1.54%、1.71%、1.28%,表明川西獐牙菜供试品溶液12 h内具有良好的稳定性。
1.3.5.7 重复性实验精密称取6份50.0 mg川西獐牙菜的干燥粉末并制备供试品溶液进行测定,结果显示獐牙菜苦苷平均含量为5.94 mg/g,RSD值为1.96%;芒果苷平均含量为45.94 mg/g,RSD值为1.58%;龙胆苦苷平均含量为102.28 mg/g,RSD值为1.35%;獐牙菜苷平均含量为2.99 mg/g,RSD值为1.99%;异荭草素平均含量为0.84 mg/g,RSD值为1.36%,结果表明运用该方法测定川西獐牙菜各成分含量具有较好的重复性。
1.3.5.8 加样回收实验精密称取6份已知含量的川西獐牙菜干燥粉末25.0 mg,每份加入等量的5份混合对照品溶液制备成供试品溶液,测定并计算各成分的加样回收率。结果显示獐牙菜苦苷平均回收率为102.06%,RSD值为1.95%;芒果苷平均回收率为101.05%,RSD值为1.30%;龙胆苦苷平均回收率为99.47%,RSD值为0.73%;獐牙菜苷平均回收率为99.67%,RSD值为2.22%;异荭草素平均回收率为99.85%,RSD值为1.77%,表明该方法回收率良好。
1.3.5.9 样品含量测定分别精密称取3份0、0.5、1.0、2.0、3.0 mmol/L SNP处理下的川西獐牙菜干燥粉末50.0 mg,测定川西獐牙菜样品中各成分含量。
1.4 统计学分析实验结果采用Graghpad Prism 8.0.2统计学软件进行分析,实验数据以均数±标准差(x±s)表示。数据符合正态分布,组间比较采用单因素方差分析。P < 0.05表示差异具有统计学意义。
2 结果与分析 2.1 SNP处理对川西獐牙菜生长的影响随着SNP处理浓度升高,川西獐牙菜的鲜质量、干质量均呈现出先增加后降低的趋势,并在1.0 mmol/L处理下达到最高,分别是0 mmol/L SNP的1.73、1.45倍。综上,低浓度SNP能够促进川西獐牙菜生长,高浓度SNP抑制川西獐牙菜生长。见图 2。

|
| 注:与0 mmol/L SNP比较,*P < 0.05,**P < 0.01。 图 2 SNP处理对川西獐牙菜生长的影响(x±s) |
在0.5、1.0、2.0 mmol/L的SNP处理时,叶绿素a、叶绿素b和类胡萝卜素含量呈现先上升后下降的趋势,并在1.0 mmol/L的SNP处理时含量达到最大,相比0 mmol/L的SNP处理分别上升了83.85%、55.65%和80.00%。当SNP浓度大于2.0 mmol/L时,3种光合色素含量又有所升高。见图 3。

|
| 注:与0 mmol/L SNP比较,*P < 0.05,**P < 0.01,***P < 0.001,****P < 0.000 1。 图 3 SNP处理对川西獐牙菜叶绿素、类胡萝卜素含量的影响(x±s) |
1.0 mmol/L的SNP处理10 d后,处理组的快速叶绿素荧光动力学曲线(O-J-I-P)趋势基本与0 mmol/L的SNP处理保持一致,说明SNP处理可能未对川西獐牙菜的PSⅡ反应中心造成破坏,见图 4。相比于0 mmol/L SNP,各个浓度SNP处理下单位反应中心捕获的用于电子传递的能量(ET0/RC)、单位反应中心捕获的用于还原QA的能量(TR0/RC)、单位反应中心吸收的光能(ABS/RC)、光合性能指数(PIABS)分别增长了14.57%、1.62%、1.16%、26.02%,而单位反应中心耗散掉的能量(DI0/RC)下降了2.30%,说明1.0 mmol/L的SNP处理增强了单位叶面积吸收光能用于电子传递的能量,同时降低了单位叶面积的热耗散,有利于提高叶片的光合性能,见图 5。SNP处理的初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)分别比0 mmol/L的SNP处理高出6.74%、4.27%、0.71%,说明SNP处理能够有效提高川西獐牙菜叶片的叶绿素荧光效能,见表 2。

|
| 图 4 SNP处理对川西獐牙菜O-J-I-P曲线的影响 |

|
| 图 5 川西獐牙菜PSⅡ比活性参数对SNP处理的响应 |

随着SNP处理浓度升高,川西獐牙菜幼苗的MDA含量呈现出先降低后升高的趋势:0.5、1.0、2.0 mmol/L的SNP处理时,MDA含量较0 mmol/L SNP处理显著降低,并且在2.0 mmol/L SNP处理时降到最低;当SNP浓度增加至3.0 mmol/L时,MDA含量略有升高,但仍低于0 mmol/L SNP处理。
随着SNP处理浓度升高,SOD活性呈现出先升高后降低的趋势,1.0 mmol/L的SNP处理时活性最高,是0 mmol/L SNP处理后活性的1.84倍;当SNP浓度超过1.0 mmol/L时,SOD活性略有下降,但仍高于0 mmol/L SNP处理。
POD和CAT活性随着SNP浓度的增加呈现出持续升高趋势。在3.0 mmol/L的SNP处理时,POD和CAT的活性达到最大值,分别是0 mmol/L SNP处理后活性的7.15、2.19倍。综上所述,在1.0、2.0、3.0 mmol/L的SNP处理时,川西獐牙菜的抗氧化酶活性较高,其中POD酶活性变化最大。见图 6。

|
| 注:与0 mmol/L SNP处理比较,*P < 0.05,**P < 0.01,***P < 0.001,****P < 0.0001。 图 6 SNP处理对川西獐牙菜MDA含量和SOD、POD、CAT酶活性的影响(x±s) |
在0.5 mmol/L的SNP处理时,川西獐牙菜幼苗中獐牙菜苦苷和异荭草素含量升高,其中异荭草素含量变化最明显,是0 mmol/L SNP处理的1.74倍;芒果苷含量略有升高,但差异无统计学意义(P>0.05)。随着SNP浓度升高,獐牙菜苦苷、芒果苷和异荭草素含量逐渐降低。在5个浓度的SNP处理下,处理组的龙胆苦苷和獐牙菜苷含量均低于0 mmol/L SNP处理。见图 7。

|
| 注:与0 mmol/L SNP处理比较,*P < 0.05,***P < 0.001,****P < 0.0001。 图 7 SNP处理对川西獐牙菜中獐牙菜苦苷、芒果苷、龙胆苦苷、獐牙菜苷、异荭草素含量的影响(x±s) |
NO作为一种关键的信号分子,能够介导多种生理功能,包括植物的生长和发育。1.0 mmol/L的SNP处理能够显著增加川西獐牙菜的鲜质量和干质量,与外源施加0.15 mmol/L的SNP增加盐胁迫下薄荷的鲜质量与干质量的研究结果相似[13]。推测可能与SNP调节细胞壁内外的β-D-葡聚糖酶积累有关[14];高浓度SNP抑制川西獐牙菜的生长,这可能与SNP能够抑制糖酵解基因参与碳水化合物代谢有关[15]。NO具有双向效应,低浓度通常会促进细胞生物量的积累,增强次生代谢产物的合成,但高浓度则可能会对植物生长产生毒性。
3.2 SNP处理对川西獐牙菜光合作用的影响叶绿素作为一种极其重要的光合色素在吸收光和传递光能中发挥至关重要的作用[16]。与0 mmol/L的SNP处理比较,1.0 mmol/L的SNP处理显著提升川西獐牙菜的叶绿素a、叶绿素b及类胡萝卜素含量。在其他研究中也观察到类似结果,如小麦幼苗和大麦幼苗在经SNP处理后叶绿素含量升高[17],推测与NO通过调节叶绿素生物合成酶的特定基因表达来影响转录或翻译有关[18]。另有研究证明SNP可以通过保护含有叶绿素的细胞器膜提高光合色素含量[17]。叶绿素荧光动力学特征是反映植物光合特征的重要参数[19]。外源NO处理能够有效提高叶片的电子传递能力,从而提高PSⅡ反应中心的活性[20]。
3.3 SNP处理对川西獐牙菜生理生化的影响SNP处理能够改变植物的代谢过程,最终导致活性氧的过量产生。超负荷的活性氧会诱发氧化应激,导致脂质过氧化和膜损伤,从而促进MDA生成[21]。在酶类抗氧化剂中,SOD酶是抗氧化的第一道防线,将氧离子(O2-)转化为过氧化氢(H2O2),随后被APX、POD和CAT所代谢。当处理川西獐牙菜的SNP浓度小于2.0 mmol/L时,MDA含量降低,SOD、POD和CAT等均表现出较高活性。随着SNP浓度增加,MDA含量升高,SOD酶活性降低,但是POD和CAT仍然保持上升趋势。而3.0 mmol/L的SNP处理后,SOD出现下降趋势,且MDA含量也较1.0 mmol/L的SNP处理时升高,这是因为SNP具有一定毒性,高浓度亚硝酸盐会对植物造成胁迫[22]。亚硝酸盐会与蛋白质的巯基与不饱和环自由基反应生成膜脂肪酸,并对细胞结构造成严重损害[23]。因此,高浓度SNP处理对植物系统具有潜在毒性,并与活性氧具有协同作用。
3.4 SNP处理对川西獐牙菜药用成分的影响川西獐牙菜中的獐牙菜苦苷、龙胆苦苷等环烯醚萜和芒果苷、异荭草素等类黄酮成分是其发挥药效的物质基础[24]。NO是启动植物次生代谢信号转导途径中的关键分子。外源施加SNP能够诱导胁迫条件下的植物大量合成次生代谢产物以应对逆境[25]。结果显示,0.5 mmol/L的SNP处理能够促进川西獐牙菜中芒果苷和异荭草素的积累,这是因为NO能够诱导苯丙氨酸代谢通路中关键酶的基因表达,从而提高相关酶活性,促进催化生成苯丙烷类次生代谢产物[26]。然而,0.5 mmol/L的SNP处理会增加獐牙菜苦苷累积,对獐牙菜苷、龙胆苦苷并无促进作用。推测低浓度SNP处理对川西獐牙菜中类黄酮成分累积的促进作用更大,而对环烯醚萜类成分累积的促进作用并不显著。而高浓度SNP处理抑制川西獐牙菜中萜类及黄酮类成分的合成,这与高浓度SNP抑制青钱柳总黄酮合成的结果一致[27]。推测高浓度亚硝酸盐胁迫导致H2O2过度累积,造成植物膜脂损伤,抑制生物合成[28]。
4 小结本研究观察SNP处理对川西獐菜的生长、生理生化指标以及药用成分积累的影响。0.5、1.0 mmol/L的SNP处理能够提高川西獐牙菜抗氧化酶活性,降低MDA含量,并对其光合作用及生长起到促进作用。在高浓度SNP处理下,SOD活性转升为降,且MDA含量转降为升,说明川西獐牙菜中产生的过量活性氧已经对其生长产生抑制作用。此外,0.5 mmol/L的SNP能够促进川西獐牙菜中獐牙菜苦苷、芒果苷和异荭草素的合成。本次实验为有关外源物质促进川西獐牙菜生长、抗氧化能力以及调控药用成分的生物合成提供了科学依据。
| [1] |
董志强, 杜少波, 陈晓文, 等. 典型藏茵陈原植物(川西獐芽菜、椭圆叶花锚、湿生扁蕾)分子水平的研究进展[J]. 安徽农学通报, 2022, 28(11): 13-15, 28. DOI:10.3969/j.issn.1007-7731.2022.11.005 |
| [2] |
赵舒媛, 胡月, 贾榕, 等. 藏药植物的分子生物学研究现状和展望[J]. 药物生物技术, 2022, 29(5): 515-521. |
| [3] |
韵海霞, 陈志. 川西獐牙菜的研究进展[J]. 华西药学杂志, 2020, 35(5): 567-571. |
| [4] |
杜茜, 张珍源, 陈西敬, 等. 川西獐牙菜药物代谢动力学和质量控制研究进展[J]. 中南药学, 2023, 21(5): 1315-1320. |
| [5] |
姚军, 郑贺云, 张翠环, 等. 外源硝普钠处理对甜瓜采后贮藏特性的影响[J]. 保鲜与加工, 2023, 23(9): 1-5. DOI:10.3969/j.issn.1009-6221.2023.09.001 |
| [6] |
王成龙, 王子然, 杨婧怡, 等. 硝普钠浸种对复合盐碱胁迫下红芸豆种子萌发和内源激素含量的影响[J]. 核农学报, 2024, 38(7): 1398-1407. |
| [7] |
任斌, 王田雨, 黄倩雪, 等. 外源NO对核桃幼苗叶多酚及其抗氧化活性的影响[J]. 西北林学院学报, 2022, 37(2): 115-120. DOI:10.3969/j.issn.1001-7461.2022.02.15 |
| [8] |
ZHENG W F, MIAO K J, ZHANG Y X, et al. Nitric oxide mediates the fungal-elicitor-enhanced biosynthesis of antioxidant polyphenols in submerged cultures of Inonotus obliquus[J]. Microbiology, 2009, 155(Pt10): 3440-3448. |
| [9] |
ZENG F S, SUN F K, LI L L, et al. Genome-scale transcriptome analysis in response to nitric oxide in birch cells: implications of the triterpene biosynthetic pathway[J]. PLoS One, 2014, 9(12): e1161-e1167. |
| [10] |
ZHENG L P, GUO Y T, WANG J W, et al. Nitric oxide potentiates oligosaccharide-induced artemisinin production in Artemisia annua hairy roots[J]. Journal of Integrative Plant Biology, 2008, 50(1): 49-55. DOI:10.1111/j.1744-7909.2007.00589.x |
| [11] |
WANG J W, ZHENG L P, WU J Y, et al. Involvement of nitric oxide in oxidative burst, phenylalanine ammonia-lyase activation and Taxol production induced by low-energy ultrasound in Taxus yunnanensis cell suspension cultures[J]. Nitric Oxide-biology and Chemistry, 2006, 15(4): 351-358. DOI:10.1016/j.niox.2006.04.261 |
| [12] |
ZHANG Y M, JIA R, HUI T Y, et al. Transcriptomic and physiological analysis of the response of Spirodela Polyrrhiza to sodium nitroprusside[J]. BMC Plant Biology, 2024, 24(1): 95. DOI:10.1186/s12870-024-04766-6 |
| [13] |
郝晓华, 雷晓琪, 高如霞. 硝普钠对盐胁迫下薄荷植株生理指标的影响[J]. 太原师范学院学报(自然科学版), 2020, 19(3): 86-90. |
| [14] |
AN L Z, LIU Y H, ZHANG M X, et al. Effects of nitric oxide on growth of maize seedling leaves in the presence or absence of ultraviolet-B radiation[J]. Journal of Plant Physiology, 2005, 162(3): 317-326. DOI:10.1016/j.jplph.2004.07.004 |
| [15] |
GU L, ZHONG X, LIAN D H, et al. Triterpenoid biosynthesis and the transcriptional response elicited by nitric oxide in submerged fermenting Ganoderma lucidum[J]. Process Biochemistry, 2017, 60(1): 19-26. |
| [16] |
闫鸿媛. 植物工厂草莓光照时长和叶面肥施用技术研究[J]. 长江蔬菜, 2024, 41(12): 65-68. |
| [17] |
ALP K, TERZI H, YILDIZ M. Proteomic and physiological analyses to elucidate nitric oxide-mediated adaptive responses of barley under cadmium stress[J]. Physiology and Molecular Biology of Plants, 2022, 28(7): 1467-1476. DOI:10.1007/s12298-022-01214-3 |
| [18] |
HAYAT S, YADAV S, WANI A S, et al. Nitric oxide effects on photosynthetic rate, growth, and antioxidant activity in tomato[J]. International Journal of Vegetable Science, 2011, 17(4): 333-348. DOI:10.1080/19315260.2011.563275 |
| [19] |
王斌, 王亚菲, 蒋媛, 等. 不同栽培模式对患枝枯病库尔勒香梨树光合及叶绿素荧光特征的影响[J]. 北方园艺, 2023, 47(8): 32-37. |
| [20] |
吴晓蕾, 焦晓聪, 刘慧兰, 等. 硝普钠对低氧胁迫下黄瓜幼苗生长、光合及叶绿素荧光参数的影响[J]. 北方园艺, 2019, 43(16): 23-28. |
| [21] |
刘贝宁, 陈发兴. 外源褪黑素对盐胁迫下念珠刺桐幼苗生理特性的缓解效应[J]. 昆明学院学报, 2024, 46(3): 99-107. |
| [22] |
李恒宇, 黄代峰, 陈晓璐, 等. 不同去蜡粉砧木南瓜品种嫁接黄瓜对硝酸盐胁迫的抗性差异[J]. 山东农业大学学报(自然科学版), 2023, 54(6): 814-819. |
| [23] |
KHEZERLUO M, HOSSEINI B, AMIRI J. Sodium nitroprusside stimulated production of tropane alkaloids and antioxidant enzymes activity in hairy root culture of Hyoscyamus reticulatus L[J]. Acta Biologica Hungarica, 2018, 69(4): 437-448. |
| [24] |
杨增亮, 刘学良, 苏媛, 等. 高效液相色谱法同时测定川西獐牙菜中10种成分的含量[J]. 西北药学杂志, 2023, 38(2): 15-19. |
| [25] |
刘慧, 吕真真, 杨文博, 等. 采后硝普钠处理对桃果实贮藏品质及抗氧化活性的影响[J]. 食品工业科技, 2023, 44(19): 374-381. |
| [26] |
宋雪, 刘思佳, 何录文, 等. 外源NO对龙胆次生代谢调控及质量形成机制研究[J]. 中草药, 2023, 54(17): 5716-5724. |
| [27] |
陈淑慧, 杭紫莹, 张昊宸, 等. 外源硝普钠对青钱柳叶片内活性物质含量和体外抗氧化能力的影响[J]. 山东林业科技, 2024, 54(2): 1-7. |
| [28] |
WU C H, TEWARI R K, HAHN E J, et al. Nitric oxide elicitation induces the accumulation of secondary metabolites and antioxidant defense in adventitious roots of Echinacea purpurea[J]. Journal of Plant Biology, 2007, 50(6): 636-643. |