 2025, Vol. 44
2025, Vol. 44文章信息
- 谷梦彤, 房城, 王旭, 闫丽, 项雨杰, 吴修红
- GU Mengtong, FANG Cheng, WANG Xu, YAN Li, XIANG Yujie, WU Xiuhong
- 中药调控脂肪酸代谢重编程干预糖尿病肾病研究进展
- Research progress of traditional Chinese medicine intervention in diabetic nephropathy based on fatty acid metabolic reprogramming
- 天津中医药大学学报, 2025, 44(4): 366-372
- Journal of Tianjin University of Traditional Chinese Medicine, 2025, 44(4): 366-372
- http://dx.doi.org/10.11656/j.issn.1673-9043.2025.04.12
-
文章历史
收稿日期: 2025-01-12

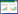



据统计,中国2型糖尿病患者中糖尿病肾病(DN)的患病率高达40%,且存在地域和性别差异[1]。DN是糖尿病加重引起的微血管并发症之一,其早期特征主要是肾小球系膜扩张、基底膜增厚和肾小球高滤过伴微量白蛋白尿,进一步发展为肾小球硬化、肾小管间质纤维化,病程发展隐匿,易错过最佳干预时期,是终末期肾病发生的主要原因[2-3]。目前DN的治疗方式主要以控制血糖、血压、血脂为主。尽管西医对其治疗手段的研究发展迅速,但仍缺乏针对性的措施以遏制其进展,故需要新的治疗方案及药物。
代谢重编程是指细胞通过调节代谢途径和代谢产物的产生与利用,来满足其一定功能与需求,主要通过影响酶蛋白关键因子的表达,以及代谢物的合成、氧化分解和转运等过程。DN患者的肾脏中存在大量脂质沉积和脂滴积累,肾脂质稳态失衡引起的脂毒性可以进一步加剧肾损伤[4]。脂肪酸作为脂质中的重要一类,细胞中的脂质代谢重编程由脂肪酸摄取、氧化和合成之间相互调节。DN中脂肪酸的代谢失调主要包括脂肪酸摄取增加、脂肪酸氧化减少、脂肪酸合成增加、脂肪酸过氧化过程。因此,分析脂肪酸代谢重编程有可能成为监测、预防和诊断DN的可行方法。研究发现中医药治疗DN具有显著疗效,与西药相比,中药治疗疾病具有多靶点、多途径、多层次、毒副作用小的特点。本研究针对脂肪酸代谢重编程与DN的内在关系、靶向脂肪酸代谢重编程治疗DN的中药进行系统综述,以期为DN的研究和治疗提供新思路。
1 脂肪酸代谢概述脂肪酸是细胞的重要能量来源和关键结构物质,还能够作为信号分子影响细胞内的信号通路传导。脂肪酸代谢是脂质代谢中的基础代谢过程,主要包括脂肪酸摄取、脂肪酸合成、脂肪酸氧化分解、脂肪酸过氧化过程。在正常情况下,脂肪酸代谢重编程受到一系列转录因子的调节,以实现系统的平衡与协调,避免脂质在细胞中的积累造成脂毒性。肾脏是合成脂肪酸的主要器官之一,脂肪酸通过相关转运蛋白,包括脂肪酸转运蛋白(FATPs)、脂肪酸结合蛋白(FABPs)、脂肪酸转位酶(CD36),进入到细胞质内参与物质合成分解代谢。脂肪酸首先活化为脂酰辅酶A(Acyl-Coa),肉碱棕榈酰转移酶(CPT1)是脂肪酸氧化(FAO)的关键酶,将Acyl-Coa转运至线粒体进行氧化分解,为机体提供能量。线粒体产生的乙酰辅酶A(Acetyl-CoA)在脂酰辅酶A合成酶(ACSL)、乙酰CoA羧化酶(ACC)、脂肪酸合成酶(FASN)、硬脂酰辅酶A去饱和酶1(SCD1)等酶的作用下合成脂肪酸,脂肪酸与体内氧化剂或代谢酶发生过氧化反应生成具有细胞毒性的脂肪酸过氧化物。目前,在组学和分子生物学等多学科研究中证实DN的发展过程中存在脂肪酸代谢重编程异常。见图 1。

|
| 注:FATPs、FABPS主要促进脂肪酸在肾小管细胞中的运输,而CD36主要处理足细胞中的脂肪酸运输。在DN情况下,体内脂肪酸在线粒体内进行氧化减少或脂肪酸合成过多超过储存在脂滴内的能力;脂肪酸过氧化经氧化应激或环氧合酶(COX)、脂氧合酶(LOX)和细胞色素P450(CYP450)转化生成脂质过氧化物,脂肪酸代谢物会在肾脏中蓄积导致足细胞凋亡或肾小管萎缩。 图 1 DN发生脂肪酸代谢异常的主要机制 |
脂肪酸是身体的重要能量来源,通过相关转运因子被吸收并运输到肾脏线粒体进行氧化。脂肪酸转运蛋白2(FATP2)是肾小管上皮细胞中主要表达的脂肪酸转运蛋白之一,可以诱导脂质代谢重编程,包括脂肪酸摄取异常,引发肾间质纤维化,与肾功能下降密切相关[5]。FABPs是参与调节细胞内脂质转运和反应的脂质伴侣,与代谢和炎症途径有关。有研究发现DN中脂肪酸相关转运蛋白的水平升高,1型DN患者的肝脏型脂肪酸结合蛋白(FABP1)和肠型脂肪酸结合蛋白(FABP2)水平显著高于健康对照组[6],FABP1因肾小管周围毛细血管血流减少引起的缺氧而释放到尿液中被识别,肾小球系膜细胞中的脂肪型脂肪酸结合蛋白(FABP4)在DN患者中呈现上调趋势[7],常作为DN的生物标志物。综上,FATP2、FABPs等转运因子通过诱导脂肪酸摄取异常引发DN中脂肪酸代谢重编程。
2.2 DN中脂肪酸氧化异常脂肪酸氧化(FAO)减少是DN中脂肪酸失衡的基础途径之一,包括如CPT1、长链脂酰辅酶A合成酶1(ACSL1)、乙酰辅酶A羧化酶2(ACC2)等表达异常,会导致脂肪酸水平在体内异常升高,从而发生肾小管上皮细胞损伤、脂质沉积、线粒体功能障碍、肾纤维化等。
通过靶向调控核心代谢酶使得FAO正常化,减少脂肪酸含量,可以缓解DN进程。Li等[8]证实,DN患者尿液中脂肪酸含量明显增加,且与肾小管间质损害程度成正比。肉碱棕榈酰转移酶1A(CPT1A)是FAO途径的重要催化剂,Xiong等[9]验证了白细胞介素-37(IL-37)通过上调CPT1A减轻DN小鼠人肾皮质近曲小管上皮细胞(HK-2)的FAO减少和肾纤维化。Wang等[10]发现ACSL1在db/db小鼠肾皮质中的表达量上调,敲除ACSL1可以抑制肿瘤坏死因子-α(TNF-α)和长链脂肪酸增加。Xu等[11]发现高糖培养HK-2中脂肪酸沉积并下调β氧化速率,破坏HK-2的表型形态,ACC2沉默处理可以加快脂肪酸β氧化速率,减少脂质沉积。此外,DN中常伴有肾小管保护因子过氧化物酶体增殖物激活受体(PPARγ)和过氧化物酶体增殖物激活受体γ共激活因子1α(PGC-1α)下调。腺苷酸活化蛋白激酶(AMPK)作为调节生物能量代谢的重要因子,可以通过调节PGC-1α来增加FAO和能量产生速率[12],减轻氧化还原失衡与糖尿病肾损伤。
综上,靶向调控脂肪酸重编程中脂肪酸氧化过程的关键酶,如ACSL1、ACC2、CPT1A及重要调节因子,如PPARγ、PGC-1α等增强FAO,抑制脂肪酸在体内蓄积,从而减少肾小管细胞损伤、脂质沉积等以改善DN。
2.3 DN中脂肪酸合成异常脂肪酸合成途径增强是DN中脂肪酸代谢异常的重要变化之一。通过影响关键酶及关键代谢产物,包括固醇调节元件结合蛋白1(SREBP-1)、碳水化合物反应元件结合蛋白(ChREBP)、FASN和ACC等,进而导致脂质沉积、胰岛素抵抗、线粒体氧化应激和炎症反应等。SREBP-1是调节脂肪酸和三酰甘油合成的靶基因,还可以作用于下游靶基因ACC和脂肪酸合酶(FAS),调控细胞内脂肪酸聚积[13],减少DN中异位脂质沉积的发生。ChREBP是高糖诱导的细胞脂肪合成的关键转录因子,被认为从脂肪酸合成、胰岛素敏感性等方面参与了DN的发病机制,Chen等[14]证明ChREBP缺失可以抑制HK-2细胞和足细胞FASN、ACC的表达,增加PPARα、CPT1A和酰基辅酶A氧化酶1(ACOX1)的表达,通过减少脂肪酸合成,降低肾脏脂质蓄积。Lu等[15]研究发现高糖刺激肾小管上皮细胞(RTECs)诱导乙酰辅酶A合成酶2(ACSS2)高表达,ACSS2可以通过调节沉默信息调节因子1(SIRT1)-ChREBP活性,抑制SIRT1表达并促进ChREBP合成脂肪酸,导致线粒体氧化应激和炎症反应,造成肾小管损伤。
因此,脂肪酸相关转运蛋白、固醇调节元件结合蛋白1c(SREBP-1c)、ChREBP的表达增加,共同导致脂肪酸合成增加,借此推测控制脂肪酸合成因子的表达,恢复正常的能量获取,可能成为DN治疗的潜在靶标。
2.4 DN中脂肪酸过氧化异常脂肪酸是体内脂质过氧化的决定因素,多不饱和脂肪酸,如花生四烯酸(AA)容易与自由基等氧化剂发生过氧化反应,COX、LOX和CYP450等酶系统也可以介导该反应发生。脂肪酸过氧化产生复杂产物,包括前列腺素、血栓素、白三烯、丙二醛等[16],具有细胞毒性和遗传毒性效应,常作为信号信使参与在DN的病理生理过程。研究证实8-异前列腺素F2α(8-iso-PGF2α)和丙二醛与肾小球滤过率的水平呈正相关关系[17]。Morita等[18]观察到前列腺素E2的代谢物四去甲前列腺素E代谢物(tetranor-PGEM)能够区分肾病的临床分期,并随着病情发展水平升高,认为tetranor-PGEM为DN新型生物标志物。酰基辅酶A合成酶长链家族成员4(ACSL4)通过平衡类花生酸生物合成来参与脂肪酸代谢,ACSL4表达上调和铁死亡加速常见于DN,抑制ACSL4可以减少脂肪酸过氧化,减弱铁死亡[19],改善肾功能。
综上,脂肪酸过氧化产物可以作为信号分子提示DN的发生,但是这一过程的分子机制现有研究不够深入,从此方面可以为防治DN提供新思路、新方法。
3 中药调控脂肪酸代谢改善DN研究进展由于中药具有多靶点的特点,作为DN治疗的主要或替代疗法,具有广阔的临床益处,越来越多的研究证明中药可以调节脂肪酸代谢重编程改善DN,主要涉及以下几个途径:干预脂肪酸摄取、干预脂肪酸氧化分解、干预脂肪酸合成、调节脂肪酸过氧化。
3.1 干预脂肪酸摄取脂肪酸通过肾小管细胞中的FATP2重吸收可以诱导线粒体损伤和核苷酸结合寡聚结构域样受体蛋白3(NLRP3)介导的炎症反应。黄芪甲苷Ⅳ是黄芪中的活性成分,Wang等[20]验证通过下调FATP2表达抑制脂肪酸转运至肾小管细胞,保护线粒体功能并抑制线粒体活性氧(mtROS)介导的NLRP3炎症激活,发挥抗氧化和抗炎作用,减轻DN中的肾小管损伤。
3.2 干预脂肪酸氧化分解肾脏中肾小管上皮细胞具有丰富的线粒体和旺盛的代谢活性,依赖FAO作为其能量来源。而DN中的肾小管内FAO过程往往受阻,线粒体超负荷导致肾小管细胞凋亡[21]。AMPK通路可以增加PGC-1α的活性,PGC-1α通过调节CPT1表达来改善脂肪酸β氧化。4-甲基厚朴酚是厚朴中提取的木质素类活性成分,可以通过激活AMPK/PGC-1α/肉碱脂酰转移酶Ⅰ肌肉亚型基因(CPT1B)介导的FAO来改善DN小鼠脂质代谢,并激活核因子E2相关因子2(Nrf2)及其下游抗氧化剂来降低肾脏氧化应激水平,间接抑制核转录因子-κB(NF-κB)途径[22]发挥对DN的肾脏保护作用。
CPT1α作为FAO途径中的主要限速酶,上调CPT1α可以增强线粒体呼吸并逆转肾小管细胞损伤。黄芩苷是中药黄芩中的黄酮苷类成分,通过直接靶向增强CPT1α表达来改善受损的FAO和线粒体呼吸,从而抑制纤维化相关蛋白的上调,减轻DN的肾纤维化程度[23]。小檗碱是黄连中的生物碱,一方面通过提高肾小管上皮细胞中CPT1A、过氧化物酶体增殖物激活受体α(PPARα)和PGC-1α的表达促进FAO,降低棕榈酸诱导的细胞内过量脂质积累,减弱细胞凋亡[24]。另外通过激活AMPK/PGC-1α促进线粒体能量稳态和FAO,抑制CD36的表达来抑制脂肪酸摄取[25],保护肾细胞免受脂毒性的损害。
PGC-1α是SIRT1下游的重要靶点之一,通过调节PGC-1α增强线粒体功能,维持肾小管上皮细胞线粒体损伤后的能量需求及代谢平衡。白藜芦醇是一种天然多酚化合物,可以通过激活AMPK/SIRT1/PGC-1α通路逆转肾脂质沉积,其中增强FAO基因PPARα和CPT1A的表达,并抑制肾细胞凋亡和氧化应激[26],从而达到对DN的缓解作用。莫诺苷是中药山茱萸中的环烯醚萜苷类成分,采用棕榈酸或高糖刺激小鼠肾小管上皮细胞建立DN体外脂肪酸代谢紊乱模型,研究表明莫诺苷可以显著上调和激活PGC-1α表达,促进胆固醇外排,抑制细胞脂质沉积,改善肾脏脂质代谢紊乱[27],预防DN发展。淫羊藿苷是淫羊藿中黄酮苷类活性成分,可以减轻脂肪酸所致肾小管上皮细胞损伤,通过诱导PGC-1α的表达激活AMPK,促进FAO,缓解脂肪酸诱导的线粒体融合-分裂失衡及抑制活性氧(ROS)生成、炎症反应和细胞凋亡[28]。
综上,AMPK、PPARα通路介导的FAO不但可以降低脂质积累,还可能降低氧化应激程度与炎症反应等。中药活性成分直接作用靶点CPT1α、PGC-1α等,主要作用在肾小管上皮细胞发挥改善DN的作用。
3.3 干预脂肪酸合成SREBP-1、ChREBP及其下游基因ACC和FASN是调控脂肪酸合成的重要转录因子。ACC可以催化过量的乙酰辅酶A转化为FAS的底物丙二酰辅酶A,丙二酰辅酶A升高抑制FAO并激活FAS活性,导致细胞中三酰甘油(TG)积累。莫诺苷是环烯醚萜苷类化合物,作用在HK-2细胞和小鼠肾脏中,可以降低NF-κB/TNF-α/SREBP-1c的表达,并抑制ACC、FAS和SCD1表达[29],抑制DN肾脏脂质沉积。肾脏脂质沉积和肾小球滤过率值低导致足细胞中的连接黏附分子样(JAML)表达较高。白藜芦醇减少JAML,增加SIRT1的表达,抑制脂肪酸合成途径中SREBP-1和ChREBP的表达,并减少高脂饮食诱导的小鼠肾脏脂质沉积与脂毒性损伤[30]。三七多糖是中药三七中的多糖成分,可以通过下调SREBP-1c以及乙酰辅酶A羧化酶α(ACCα)的水平,调节脂质代谢紊乱,降低血液中脂肪含量[31]。贞清方由女贞子、墨旱莲、山药3味药物组成,具有补肾养阴、活血化瘀、通络利尿的功效,可以抑制肾皮质中SREBP-1c及其靶基因ACC与FAS的过表达,减少脂肪酸合成,抑制肾皮质中转化生长因子-β1(TGF-β1)的表达,从而减少纤连蛋白和细胞外基质扩张[32],改善DN的肾功能。
此外,中药可以多途径联合脂肪酸合成及FAO过程中的靶点调节脂肪酸代谢。PPARα在肾脏中表达丰富,主要调节下游靶基因ACOX1抑制FAO,PPARs在PGC-1α的协同作用下促进CPT1A的蛋白表达。花青素干预后,近曲小管上皮细胞内脂滴减少,降低SREBP-1、FASN及ACC的表达,升高PPARα、CPT1及ACOX1的表达,改善糖尿病小鼠肾组织的异常脂质沉积[33]。百令胶囊为冬虫夏草发酵菌粉,具有补肺肾、益精气的功效。Zhang等[34]通过激活DN中PPARα、ACOX1和硬脂酰辅酶A去饱和酶(SCD)的表达,抑制FASN的表达,促进FAO,减少肾脏脂质积聚,延缓DN进展。
综上,中药不仅通过抑制SREBP-1c、ChREBP介导的脂肪酸合成,作用下游靶点包括ACC、FASN、SCD1等,还通过激活PPARα信号通路促进FAO,以多靶点、多调节途径的方式治疗DN。
3.4 调节脂肪酸过氧化中药可以调节脂肪酸过氧化发挥对DN的保护作用,花生四烯酸代谢(AA代谢)中环氧二十碳三烯酸(EETs)和PPARγ通路存在密切联系,抑制PPARγ的表达会减弱EETs的抗炎作用。藏红花素是中药番红花的主要酯类成分,可以降低AA、11,12-EET的水平,激活细胞色素P450 4A11(CYP4A11)/PPARγ通路,抑制转化生长因子-β(TGF-β)/斯马德蛋白(Smad)通路[35],对DN小鼠的治疗起到关键作用。青藤碱作用于亚油酸代谢、AA代谢等代谢途径[36],降低细胞色素P450 1A1(CYP1A1)表达,并减少肾脏中白三烯B4水平。地黄叶总苷可以降低AA代谢通路中前列腺素H3(PGH3)水平,影响TGF-β/Smad通路[37],对db/db小鼠的早期肾损伤起到保护作用。黄芪和党参可以通过回调AA代谢,增加19(S)-羟基二十碳四烯酸[19(S)-HETE]含量,抑制前列腺素过氧化物合酶2的表达[38],调节肾脏血流动力学,改善肾小球和肾小管的代谢紊乱,减轻肾功能损害。
玉米须多糖可以调节AA代谢、α-亚麻酸代谢、不饱和脂肪生物合成等代谢通路,回调亚麻酸、EETs、血栓素B2(TXB2)等多种代谢物水平缓解DN[39]。丹参酚酸B和丹参酮ⅡA是丹参重要的活性成分,能够回调2, 3-Dinor-TXB2,抑制血栓素A2(TXA2)合成,调节磷脂酰肌醇3-激酶(PI3K)/蛋白激酶B(Akt)/NF-κB信号通路。此外,中药丹参也可以通过调节饱和脂肪酸的生物合成、AA代谢等代谢通路,抑制高糖诱导肾小球系膜细胞中wnt4、β-catenin和TGF-β的相对表达水平[40-41],改善早期DN大鼠糖脂紊乱、肝肾损伤,抵抗肾脏炎症。
肾炎康复片具有益气养阴、健脾补肾、清解毒邪之效,Wang等[42]发现不饱和脂肪酸生物合成代谢是与DN非常相关的途径之一,主要由亚油酸、油酸、AA等物质参与,认为调节脂肪酸的含量可以减缓胰岛素抵抗对人体的损害,恢复机体的代谢平衡。竹节人参具有散瘀止血、补虚强壮的功效,Wang等[43]研究发现竹节人参可以调节不饱和脂肪酸的生物合成,减少油酸和硬脂酸代谢产物,调节B细胞淋巴瘤2蛋白(Bcl-2)/半胱天冬酶3(Caspase-3)信号通路,抑制肾细胞凋亡。
综上,中药通过调节脂肪酸过氧化物水平及CYP450、COX酶系统,平衡体内脂肪酸过氧化物水平,影响炎症通路,调节肾血流动力学,改善糖脂代谢紊乱,缓解胰岛素抵抗抑制肾细胞凋亡等,改善DN。见开放科学(资源服务)标识码(OSID)。
4 总结与展望综上所述,脂肪酸代谢重编程中的关键因子与DN之间存在密切的关联性,是防治DN的广阔前景。DN中过量的脂肪酸会溢出到肾脏中,造成异位脂质积累,诱发脂毒性,加重肾脏损伤。多项研究从调控脂肪酸代谢重编程角度,包括脂肪酸摄取、脂肪酸氧化分解、脂肪酸合成、脂肪酸过氧化途径,阐述了中医药防治DN的机制,具有一定的有效性。与西药相比,中药作用途径复杂,可以多条路径协同调节改善DN,如莫诺苷参与脂肪酸氧化与合成两条途径。中药调控脂肪酸代谢重编程还涉及改善脂质沉积、降低炎症反应、改善线粒体功能障碍、改善肾脏固有细胞及胰岛细胞凋亡等多种机制。
目前,中药对脂肪酸代谢重编程的研究仍然存在一些不足,研究者们多通过肾脏固有细胞及动物实验验证中药调控脂肪酸代谢重编程的相关机制,还需要进一步临床试验验证是否直接靶向定位肾脏病变部位发挥作用改善DN;中药对干预脂肪酸摄取过程中跨膜转运蛋白靶点,如FABPs、FATPs的相关研究较少,今后需要更多验证;DN中脂肪酸蓄积引发脂毒性,研究者们重点从温补脾肾、活血祛瘀化浊、除湿祛毒功效为主的中药进行挖掘,化浊降脂改善肾脂质沉积。相信随着脂肪酸代谢重编程与DN相关研究的不断深入,调控脂肪酸代谢重编程防治DN的理念可以为探讨中医药在防治DN的基础研究与临床应用方面提供新思路、新依据。
| [1] |
ZHANG X X, KONG J, YUN K. Prevalence of diabetic nephropathy among patients with type 2 diabetes mellitus in China: a meta-analysis of observational studies[J]. Journal of Diabetes Research, 2020, 20(11): 2315-2327. |
| [2] |
GUPTA S, DOMINGUEZ M, GOLESTANEH L. Diabetic kidney disease an update[J]. Medical Clinics of North America, 2023, 107(4): 689-705. DOI:10.1016/j.mcna.2023.03.004 |
| [3] |
KOYE D N, MAGLIANO D J, NELSON R G, et al. The global epidemiology of diabetes and kidney disease[J]. Advances in Chronic Kidney Disease, 2018, 25(2): 121-132. DOI:10.1053/j.ackd.2017.10.011 |
| [4] |
WANG Y, LIU T T, WU Y, et al. Lipid homeostasis in diabetic kidney disease[J]. International Journal of Biological Sciences, 2024, 20(10): 3710-3724. DOI:10.7150/ijbs.95216 |
| [5] |
CHEN Y T, YAN Q, LV M Y, et al. Involvement of FATP2-mediated tubular lipid metabolic reprogramming in renal fibrogenesis[J]. Cell Death & Disease, 2020, 11(11): 994-1004. |
| [6] |
ABO EL-ASRAR M, ISMAIL E A R, ELNHRAWY A M, et al. Fatty acid binding protein 1(FABP1) and fatty acid binding protein 2(FABP2) as a link between diabetic nephropathy and subclinical atherosclerosis in children and adolescents with type 1 diabetes[J]. Journal of Diabetes and Its Complications, 2023, 37(3): 1084-1104. |
| [7] |
YAO F, LI Z, EHARA T, et al. Fatty Acid-Binding Protein 4 mediates apoptosis via endoplasmic reticulum stress in mesangial cells of diabetic nephropathy[J]. Molecular and Cellular Endocrinology, 2015, 411(2): 232-242. |
| [8] |
LI L, WANG C S, YANG H L, et al. Metabolomics reveal mitochondrial and fatty acid metabolism disorders that contribute to the development of DKD in T2DM patients[J]. Molecular BioSystems, 2017, 13(11): 2392-2400. DOI:10.1039/C7MB00167C |
| [9] |
XIONG L, HE T, LIU C, et al. IL-37 ameliorates renal fibrosis by restoring CPT1A-mediated fatty acid oxidation in diabetic kidney disease[J]. Kidney Diseases(Basel, Switzerland), 2023, 9(2): 104-117. |
| [10] |
WANG C H, SURBHI, GORAYA S, et al. Fatty acids and inflammatory stimuli induce expression of long-chain acyl-CoA synthetase 1 to promote lipid remodeling in diabetic kidney disease[J]. Journal of Biological Chemistry, 2024, 300(1): 105-115. |
| [11] |
XU Y, HUANG J, XIN W, et al. Lipid accumulation is ahead of epithelial-to-mesenchymal transition and therapeutic intervention by acetyl-CoA carboxylase 2 silence in diabetic nephropathy[J]. Metabolism, 2014, 63(5): 716-726. DOI:10.1016/j.metabol.2014.02.010 |
| [12] |
NAGAI Y, MATOBA K, TAKEDA Y, et al. Rho-associated, coiled-coil-containing protein kinase 1 regulates development of diabetic kidney disease via modulation of fatty acid metabolism[J]. Kidney International, 2022, 102(3): 536-545. DOI:10.1016/j.kint.2022.04.021sn.1007-3728.2016.11.007ry.action?journalTitle=思想战线&year=2019&firstpage=50&issue=06 |
| [13] |
DIHINGIA A, BORDOLOI J, DUTTA P, et al. Hexane-Isopropanolic Extract of Tungrymbai, a North-East Indian fermented soybean food prevents hepatic steatosis via regulating AMPK-mediated SREBP/FAS/ACC/HMGCR and PPARα/CPT1A/UCP2 pathways[J]. Scientific Reports, 2018, 8(1): 100-121. DOI:10.1038/s41598-017-18531-3 |
| [14] |
CHEN N, MU L, YANG Z F, et al. Carbohydrate response element-binding protein regulates lipid metabolism via mTOR complex1 in diabetic nephropathy[J]. Journal of Cellular Physiology, 2021, 236(1): 625-640. DOI:10.1002/jcp.29890 |
| [15] |
LU J, LI X Q, CHEN P P, et al. Acetyl-CoA synthetase 2 promotes diabetic renal tubular injury in mice by rewiring fatty acid metabolism through SIRT1/ChREBP pathway[J]. Acta Pharmacologica Sinica, 2024, 45(2): 366-377. DOI:10.1038/s41401-023-01160-0 |
| [16] |
BOGIE J F J, HAIDAR M, KOOIJ G, et al. Fatty acid metabolism in the progression and resolution of CNS disorders[J]. Advanced Drug Delivery Reviews, 2020, 159(1): 198-213. |
| [17] |
SAURIASARI R, ANDRAJATI R, AZIZAHWATI, et al. Marker of lipid peroxidation related to diabetic nephropathy in Indonesian type 2 diabetes mellitus patients[J]. Diabetes Research and Clinical Practice, 2015, 108(1): 193-200. DOI:10.1016/j.diabres.2014.12.016 |
| [18] |
MORITA Y, KURANO M, SAKAI E, et al. Simultaneous analyses of urinary eicosanoids and related mediators identified tetranor-prostaglandin E metabolite as a novel biomarker of diabetic nephropathy[J]. Journal of Lipid Research, 2021, 62(5): 100-120. |
| [19] |
DING K Y, LIU C B, LI L, et al. Acyl-CoA synthase ACSL4: an essential target in ferroptosis and fatty acid metabolism[J]. Chinese Medical Journal, 2023, 136(21): 2521-2537. |
| [20] |
WANG J, WANG L C, FENG X X, et al. Astragaloside Ⅳ attenuates fatty acid-induced renal tubular injury in diabetic kidney disease by inhibiting fatty acid transport protein-2[J]. Phytomedicine, 2024, 134(12): 1559-1591. |
| [21] |
KRUGER C, NGUYEN T T, BREAUX C, et al. Proximal tubular cell-specific ablation of carnitine acetyltransferase causes tubular disease and secondary glomerulosclerosis[J]. Diabetes, 2019, 68(4): 819-831. DOI:10.2337/db18-0090 |
| [22] |
MA T J, ZHENG Z Y, GUO H, et al. 4-O-methylhonokiol ameliorates type 2 diabetes-induced nephropathy in mice likely by activation of AMPK-mediated fatty acid oxidation and Nrf2-mediated anti-oxidative stress[J]. Toxicology and Applied Pharmacology, 2019, 370(1): 93-105. |
| [23] |
HU H T, LI W W, HAO Y Q, et al. Baicalin ameliorates renal fibrosis by upregulating CPT1α-mediated fatty acid oxidation in diabetic kidney disease[J]. Phytomedicine, 2024, 122(4): 1551-1562. |
| [24] |
SUN J Y, CHEN X M, LIU T, et al. Berberine protects against palmitate-induced apoptosis in tubular epithelial cells by promoting fatty acid oxidation[J]. Medical Science Monitor, 2018, 24(10): 1484-1492. |
| [25] |
QIN X, JIANG M, ZHAO Y, et al. Berberine protects against diabetic kidney disease via promoting PGC-1α-regulated mitochondrial energy homeostasis[J]. British Journal of Pharmacology, 2020, 177(16): 3646-3661. DOI:10.1111/bph.14935 |
| [26] |
CHANG C C, CHANG C Y, WU Y T, et al. Resveratrol retards progression of diabetic nephropathy through modulations of oxidative stress, proinflammatory cytokines, and AMP-activated protein kinase[J]. Journal of Biomedical Science, 2011, 18(1): 47-62. DOI:10.1186/1423-0127-18-47 |
| [27] |
GAO J W, LIU P, SHEN Z R, et al. Morroniside promotes PGC-1 α-mediated cholesterol efflux in sodium palmitate or high glucose-induced mouse renal tubular epithelial cells[J]. BioMed Research International, 2021, 109(18): 9942-9952. |
| [28] |
吴艺璇, 彭维艳, 刘露, 等. 淫羊藿苷通过AMPK减轻脂肪酸诱导的肾小管上皮细胞线粒体损伤[J]. 中国病理生理杂志, 2022, 38(1): 113-121. DOI:10.3969/j.issn.1000-4718.2022.01.015 |
| [29] |
ZHU W H, CHEN M, WANG Y, et al. Regulation of renal lipid deposition in diabetic nephropathy on morroniside via inhibition of NF-κB/TNF-a/SREBP1c signaling pathway[J]. Chemico-Biological Interactions, 2023, 385(11): 1107-1121. |
| [30] |
GU W, WANG X, ZHAO H F, et al. Resveratrol ameliorates diabetic kidney injury by reducing lipotoxicity and modulates expression of components of the junctional adhesion molecule-like/sirtuin 1 lipid metabolism pathway[J]. European Journal of Pharmacology, 2022, 918(14): 1747-1756. |
| [31] |
李易, 叶曦. 三七多糖对糖尿病肾病大鼠炎症反应及脂质代谢调节作用的实验研究[J]. 中国中医药科技, 2018, 25(1): 43-47. |
| [32] |
WEN X Y, ZENG Y L, LIU L F, et al. Zhenqing recipe alleviates diabetic nephropathy in experimental type 2 diabetic rats through suppression of SREBP-1c[J]. Journal of Ethnopharmacology, 2012, 142(1): 144-150. DOI:10.1016/j.jep.2012.04.028 |
| [33] |
杜春阳, 姚芳, 任韫卓, 等. 葡萄籽原花青素对db/db小鼠肾组织脂质沉积的影响[J]. 中国细胞生物学学报, 2017, 39(3): 288-295. |
| [34] |
ZHANG Q, XIAO X H, LI M, et al. Bailing capsule(Cordyceps sinensis) ameliorates renal triglyceride accumulation through the PPARα pathway in diabetic rats[J]. Frontiers in Pharmacology, 2022, 13(12): 915-925. |
| [35] |
CHEN W, SU J H, LIU Y B, et al. Crocin ameliorates diabetic nephropathy through regulating metabolism, CYP4A11/PPARγ, and TGF-β/smad pathways in mice[J]. Current Drug Metabolism, 2023, 24(10): 709-722. DOI:10.2174/0113892002257928231031113337 |
| [36] |
LI Y, WANG L, ZHANG J M, et al. Integrated multi-omics and bioinformatic methods to reveal the mechanisms of sinomenine against diabetic nephropathy[J]. BMC Complementary Medicine and Therapies, 2023, 23(1): 287-298. DOI:10.1186/s12906-023-04119-0 |
| [37] |
XU Z, DAI X X, ZHANG Q Y, et al. Protective effects and mechanisms of Rehmannia glutinosa leaves total glycoside on early kidney injury in db/db mice[J]. Biomedicine & Pharmacotherapy, 2020, 125(10): 1099-1126. |
| [38] |
SHEN Z, CUI T, LIU Y, et al. Astragalus membranaceus and Salvia miltiorrhiza ameliorate diabetic kidney disease via the "gut-kidney axis"[J]. Phytomedicine, 2023, 121(2): 155-169. |
| [39] |
DONG W T, ZHAO Y Y, LI X W, et al. Corn silk polysaccharides attenuate diabetic nephropathy through restoration of the gut microbial ecosystem and metabolic homeostasis[J]. Frontiers in Endocrinology, 2023, 14(6): 1232-1243. |
| [40] |
XIANG X, CAI H D, SU S L, et al. Salvia miltiorrhiza protects against diabetic nephropathy through metabolome regulation and Wnt/β-catenin and TGF-β signaling inhibition[J]. Pharmacological Research, 2019, 139(1): 26-40. |
| [41] |
XU Z, CAI K, SU S L, et al. Salvianolic acid B and tanshinone ⅡA synergistically improve early diabetic nephropathy through regulating PI3K/Akt/NF-κB signaling pathway[J]. Journal of Ethnopharmacology, 2024, 319(7): 1173-1185. |
| [42] |
WANG X L, HE Q Y, CHEN Q, et al. Network pharmacology combined with metabolomics to study the mechanism of Shenyan Kangfu Tablets in the treatment of diabetic nephropathy[J]. Journal of Ethnopharmacology, 2021, 270(10): 1138-1147. |
| [43] |
WANG T T, HUANG X T, ZHAI K F, et al. Integrating metabolomics and network pharmacology to investigate Panax japonicus prevents kidney injury in HFD/STZ-induced diabetic mice[J]. Journal of Ethnopharmacology, 2023, 303(9): 1158-1179. |