 2026, Vol. 45
2026, Vol. 45文章信息
- 张睿瑄, 刘海朝, 金昱彤, 石悦暄, 王继达, 李梦琪, 边育红
- ZHANG Ruixuan, LIU Haizhao, JIN Yutong, SHI Yuexuan, WANG Jida, LI Mengqi, BIAN Yuhong
- 调控肠道平滑肌中线粒体改善阳虚型便秘的研究进展
- The research progress on regulating mitochondria in intestinal smooth muscle to improve yang deficiency-related constipation
- 天津中医药大学学报, 2026, 45(4): 479-492
- Journal of Tianjin University of Traditional Chinese Medicine, 2026, 45(4): 479-492
- http://dx.doi.org/10.11656/j.issn.1673-9043.2026.04.14
-
文章历史
收稿日期: 2026-01-25

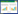
引用本文 |



2. 天津市第一中心医院, 天津 300192;
3. 天津市中西医结合危重病与急救研究所, 天津 300192;
4. 天津中医药大学第一附属医院, 天津 300381;
5. 中医国家临床医学研究中心, 天津 300381
2. Tianjin First Central Hospital, Tianjin 300192, China;
3. Tianjin Institute of Integrated Traditional Chinese and Western Medicine for Critical Care and Emergency Medicine, Tianjin 300192, China;
4. First Teaching Hospital of Tianjin University of Traditional Chinese Medicine, Tianjin 300381, China;
5. National Clinical Research Center for Chinese Medicine, Tianjin 300381, China
慢性便秘,是一种常见且影响深远的功能性胃肠疾病,其特征包括排便困难、排便次数减少、粪便干硬、有排便不尽感等,病程通常持续6个月以上[1]。慢性便秘的全球流行率为15%[2],老年人群的发病率更高,可达20%甚至70%[3-4]。目前,便秘的治疗方法包括调节饮食、增加运动、使用非处方药物等,临床上常用促动力药如多潘立酮、莫沙必利和抗痉挛药如匹维溴铵等调节胃肠动力,虽然在一定程度上可以缓解症状,但是长期使用存在诸多不良反应,如心律失常、头晕乏力、肝肾负担加重等,且疗效不稳定、复发率高[5-6]。鉴于慢性便秘的发病机制复杂、复发率高及治疗反应差等特点,它已经成为严重影响公众健康与生活质量的重要慢性疾病。相较于西医治疗手段,中医药因其多靶点作用机制、较少的不良反应、良好的安全性与耐受性以及稳定持久的疗效,在便秘治疗中展现出独特且显著的临床优势。
慢性便秘在中医里属于“脾约”“秘结”范畴[7]。参照中华中医药学会脾胃病分会的慢性便秘中医诊疗共识意见及国家中医药管理局慢传输型便秘中医临床路径、诊疗方案,将慢性便秘分为5个证型:肠道实热证、肝脾不调证、肺脾气虚证、肝肾阴虚证与脾肾阳虚证[8]。1项对300例慢性便秘患者的研究显示,阳虚型便秘在慢性便秘中占比最高,达到50.34%。研究还采用了Wexner量表对各证型的症状严重度进行了量化评估,结果显示阳虚型便秘的症状最为严重[9]。该证患者多表现为畏寒肢冷、乏力倦怠、排便无力、便质偏干等症状,提示其病机与机体代谢及能量供应不足密切相关。近年来,针对阳虚型便秘发病机制及干预途径的研究逐渐成为慢性便秘领域的研究热点。
在慢性便秘患者中,多见肠道蠕动功能下降、肠道平滑肌收缩力减弱、传导速度降低[10]。肠道平滑肌作为实现胃肠蠕动和推进功能的关键执行者,是消化系统的重要组成部分,其节律性收缩直接调控消化、吸收和排泄功能。当肠道平滑肌功能受损时,肠道蠕动力下降,导致慢性便秘的发生和加重。研究发现,阳虚型便秘患者常伴有肠道平滑肌收缩力不足与运动协调障碍,提示平滑肌功能异常可能是阳虚型便秘的重要病理环节。
平滑肌的收缩活动高度依赖线粒体提供的能量支持。线粒体作为细胞的能量工厂,担负着提供能量、调节钙离子平衡等维持细胞生理功能的重要任务。线粒体功能受损,肠道平滑肌的收缩与舒张能力将受到影响,从而引发肠道传输功能的紊乱[11]。而在中医理论中,“阳虚”状态本质上反映的是机体代谢活性下降、能量生成不足的生理失衡,与西医中线粒体功能障碍导致的能量代谢失衡具有高度一致性。阳虚机体代谢率低、产热不足、能量供应匮乏,与线粒体功能障碍所呈现的“低能耗—低功能”特征在病理生理层面高度吻合。因此,从“阳虚—线粒体功能障碍”这一视角出发,推测阳虚型便秘可能是肠道平滑肌线粒体功能障碍的宏观表现。
近年来,越来越多研究证明中医疗法能够通过调节肠道平滑肌中的线粒体功能改善肠道病理状态,从而对胃肠动力障碍类疾病发挥治疗作用[12-14]。提示从调控肠道平滑肌细胞中的线粒体入手恢复肠道平滑肌功能或将成为改善阳虚型便秘的新切入点。本研究将围绕“通过调控肠道平滑肌线粒体功能改善阳虚型便秘”这一新视角进行系统综述,探讨其潜在机制与研究前景,为中西医结合治疗阳虚型便秘提供理论依据。
1 线粒体在肠道平滑肌中的生理作用线粒体是调控细胞命运的关键细胞器,由双层膜构成。外膜含孔蛋白通道,内膜高度折叠形成嵴,嵴中富含电子传递链(ETC)和三磷酸腺苷合酶,是能量代谢的核心区域。基质内含三羧酸循环酶、线粒体脱氧核糖核酸(DNA)与核糖体,具备部分自主合成功能[15]。线粒体调控真核细胞代谢稳态、钙信号、氧化还原平衡,这些功能依赖于其结构完整性、代谢酶系统、遗传信息及动态行为的协同调节。线粒体通过融合、分裂和自噬维持自身稳态。线粒体结构变化常预示功能状态的改变,如高代谢时嵴密度增加、融合增强以提升三磷酸腺苷(ATP)合成效率;嵴断裂、膜电位丧失等改变则可以诱发细胞功能障碍甚至凋亡[16]。
肠道平滑肌细胞在多种影响下可逆地在收缩表型与非收缩表型之间转换,包括来自线粒体的许多影响[11]。具体而言,肠道平滑肌细胞的动力特性受到细胞膜兴奋性、电-机械耦联机制及钙信号通路的协调调控[17],而这些功能高度依赖线粒体供能系统的完整性与效率。线粒体作为真核细胞的能量代谢中心,通过氧化磷酸化(OXPHOS)途径为细胞活动提供ATP,参与细胞内钙稳态维持、活性氧(ROS)清除、细胞凋亡及结构自稳机制的调节[18],是保证肠道平滑肌功能稳态的基础。肠道平滑肌细胞中的线粒体功能受损会导致肠道平滑肌收缩频率紊乱,在消化系统中引发食物滞留或过早通过等动力障碍,引起功能性胃肠病、慢性便秘等疾病。随着对功能性胃肠动力障碍等疾病机制研究的深入,线粒体在肠道平滑肌功能的研究中逐渐成为一个重要切入点。
1.1 OXPHOS进程的推动线粒体是细胞能量的工厂,通过OXPHOS过程生成ATP,为细胞提供必需的能量,这一过程依赖于ETC和ATP合成酶的协同作用。ATP的生成始于线粒体内膜的电子传递链,其中4个主要的呼吸链蛋白复合物(Ⅰ-Ⅳ)在电子传递过程中逐步将电子从底物传递到最终的电子受体——氧分子,同时释放出能量[19]。这一过程中,复合物Ⅰ和Ⅲ泵送质子(H+)使其通过内膜传递到膜间隙,形成跨膜质子梯度,即电化学梯度。而复合物Ⅳ则促使氧与质子结合生成水。质子梯度的建立为ATP合成提供了驱动力,在线粒体内膜上的ATP合成酶则通过质子流动驱动ATP合成。在ATP合成酶的F0部分,质子通过ATP合成酶的通道返回基质;与此同时,F1部分利用质子的流动,驱动二磷酸腺苷(ADP)与无机磷酸(Pi)结合生成ATP。而线粒体内的氧作为终极电子受体,参与电子的连续流动和质子梯度的形成,并与质子结合生成水,维持ATP合成过程的进行[20-21]。此过程(图 1)不仅是细胞能量代谢的核心,也是维持细胞生命活动的基础。此外,三羧酸循环在线粒体基质中进行,乙酰辅酶A通过氧化产生还原型辅酶:还原型烟酰胺腺嘌呤二核苷酸(NADH)和还原型黄素腺嘌呤二核苷酸(FADH2),NADH和FADH2通过复合物Ⅰ和Ⅱ向电子传递链提供电子,并释放CO2,推动质子泵送并支持ATP合成[22]。

|
| 图 1 线粒体内电子传递链工作与ATP产生过程 |
ATP不足会影响肠道平滑肌肌球蛋白的三磷酸腺苷酶(ATPase)活性,其与肌动蛋白结合并完成收缩周期的能力下降,进而削弱每次收缩的力学输出。研究证实,ATP缺乏可以降低肌球蛋白Mg2+-ATPase活性,并显著减慢肌丝滑动速率[23]。此外,ATP生成减少还会直接影响离子泵活动、钙泵回收、肌动蛋白解聚等关键耗能过程,多层次地破坏肠道平滑肌电活动、钙动态与机械输出,从而造成肠道平滑肌蠕动无力、推进力下降、肌张力失控等病理现象。因此,肠道平滑肌内线粒体维持OXPHOS正常水平的生理作用是保证肠道平滑肌运动功能的基础。
1.2 钙稳态的维持线粒体主要通过钙离子的摄取、储存和释放来维持细胞内的钙平衡[24]。线粒体钙单向转运体(MCU)是负责将钙离子从胞质转运到线粒体基质的关键通道,其活性受到线粒体膜电位和复合体Ⅰ功能的调控。钙离子进入线粒体基质后会与磷酸盐结合形成不溶性沉淀,从而实现短期储存;同时,线粒体也会通过Na+/Ca2+交换体(NCLX)将钙离子从基质释放回胞质,维持细胞内外的钙浓度平衡。正常情况下,线粒体通过摄取胞质Ca2+缓冲细胞内钙浓度波动,从而维持平滑肌细胞电-机械耦合机制[25]。线粒体功能障碍时,钠钾泵(Na+/K+-ATPase)和钙泵的功能受限,导致胞内钙离子无法有效回收,造成细胞内钙超载,诱发肌球蛋白轻链磷酸化异常,破坏兴奋-收缩耦合节律[26-27]。研究表明,在肠道平滑肌细胞暴露于γ射线应激后,也可以观察到Ca2+失调与线粒体凋亡协同出现[28]。此外,ATP也是K+通道(如ATP敏感型钾通道,KATP)的直接调节因子。当线粒体功能出现障碍,ATP浓度下降,KATP通道开放,会导致膜超极化、电活动减弱、Ca2+通道激活障碍,从而减少胞内Ca2+浓度上升和收缩反应[29]。KATP通道活性升高与收缩力下降密切相关这一事实在糖尿病胃平滑肌中已被证实[30]。由此可知,肠道平滑肌兴奋-收缩耦合的正常功能离不开其内部线粒体对钙稳态的维持。
1.3 ROS的双重调节线粒体是细胞内主要的ROS生成场所之一,其ETC中的复合物Ⅰ和Ⅲ在OXPHOS过程中会发生电子泄漏,生成超氧阴离子。在正常生理状态下,ROS作为次级信使,参与调节细胞增殖、分化等多种重要生物学过程[19]。为了防止ROS对细胞造成损害,线粒体会通过超氧化物歧化酶(SOD2)将超氧阴离子转化为过氧化氢(H2O2),然后进一步通过谷胱甘肽过氧化物酶(GPx)和过氧化氢酶(CAT)将其清除,从而维持细胞内氧化还原平衡[31-32]。已有研究表明,长寿命物种的线粒体通常具有较低的ROS泄漏率和更强的抗氧化防御能力。例如,裸鼹鼠的线粒体膜中不饱和脂肪酸的含量较低,这一特性显著降低了其膜脂质的过氧化敏感性[33]。而当线粒体受损或功能障碍时(如线粒体基质内钙过载等),OXPHOS效率降低,ROS产生急剧增加。ROS的过量积聚会通过多种机制加剧平滑肌的结构性与功能性损伤。一方面,ROS可以通过氧化修饰细胞膜离子通道与受体蛋白,降低其激活效率,尤其是电压门控钙通道和肌动蛋白调控蛋白,从而削弱钙信号的有效性,造成肌丝滑动减慢[34]。另一方面,ROS可以直接氧化肌球蛋白与肌动蛋白的巯基位点,导致肌原纤维交联、刚性增强和动力学失调,进而降低肌细胞的机械伸缩效率[35]。同时,氧化损伤还可以通过氧化DNA和脂质激活促炎和凋亡信号使细胞无法维持合适的膜电位与代谢稳态,出现肌细胞损伤甚至丧失节律性收缩功能,发生凋亡前期的功能减退,最终使得细胞丧失收缩节律与应激反应能力[36]。因此,肠道平滑肌功能与结构的完整性及肠道平滑肌中线粒体对ROS的平衡能力密切相关。
1.4 细胞凋亡的调控线粒体是细胞凋亡的关键调控中心,其功能障碍时可以通过细胞色素C释放增加、Caspase通路异常激活等机制,促进肠道平滑肌细胞的凋亡,影响肌层结构与功能[37]。细胞凋亡通常始于线粒体膜电位(ΔΨm)的崩溃,导致细胞色素C从线粒体释放到胞质。B细胞淋巴瘤-2(Bcl-2)家族蛋白调控这一过程:抗凋亡蛋白Bcl-2抑制细胞色素C的释放,而促凋亡蛋白Bcl-2相关X蛋白(Bax)和Bcl-2同源拮抗剂(Bak)通过寡聚化形成孔道,增加线粒体外膜的通透性,从而促进细胞色素C的释放[38]。最新研究表明,Bax/Bak寡聚化不仅增强了线粒体外膜的通透性,还激活内膜蛋白酶OMA1,后者通过切割视神经萎缩蛋白1(OPA1)蛋白改变线粒体嵴结构,进一步促进细胞色素C的释放。
在线粒体功能障碍的情况下,Bax和Bak等促凋亡蛋白通过增加外膜通透性,激活凋亡信号,诱导Caspase-9与Caspase-3级联激活,导致肠道平滑肌细胞程序性死亡,削弱肌层厚度与收缩张力。同时,肌动蛋白骨架系统解聚和细胞间连接断裂,破坏肌纤维排列与同步性,影响推进性蠕动的正常进程[39]。研究还发现,线粒体介导的凋亡常伴随ATP耗竭、ROS积聚和离子通道失控等,形成代谢衰竭与结构崩解的恶性循环。在术后肠麻痹或放射损伤模型中,线粒体凋亡信号异常激活与平滑肌功能障碍高度相关,抑制凋亡相关蛋白(如Bax)或增强抗凋亡因子(如Bcl-2)表达可以逆转这一过程,进一步验证了线粒体凋亡机制在肠道平滑肌功能中的决定性作用[40-41]。
因此,肠道平滑肌组织结构完整性与动力功能正常运转离不开肠道平滑肌内线粒体所调控的细胞凋亡进程有序进行。
2 线粒体功能障碍影响肠道平滑肌功能的机制与循环 2.1 线粒体功能障碍介导的肠道平滑肌功能失调通路由前文可知,肠道平滑肌的功能依赖于其中线粒体正常生理功能的支持。研究表明,多个信号通路和调控因子调节线粒体的ATP生成、钙稳态、氧化还原反应与动力学行为等,进而控制肠道平滑肌的收缩能力、节律性和结构稳定性。目前,已经发现多个与平滑肌功能障碍相关的线粒体功能靶点与通路。
2.1.1 ATP生成异常mTORC2-Akt-GLUT4通路调节葡萄糖代谢与ATP生成:在糖尿病大鼠胃平滑肌中,Rictor/mTORC2/Akt/GLUT4通路表达异常,影响葡萄糖摄取和氧化代谢。该通路调控线粒体糖酵解与OXPHOS过程,在糖尿病胃动力障碍中出现失调,表现为ATP的合成下降、收缩频率以及幅度减弱[42-43]。
ANT-ATP转运通路维持线粒体输出与细胞能量状态:腺苷酸转运蛋白(ANT)负责线粒体内外ATP/ADP交换,是维持细胞能量平衡的重要通道。研究显示,线粒体融合蛋白1(MFN1)下调可以促进ANT活性增加、线粒体膜电位增强,但同时诱导Ca2+失稳和代谢失衡,提示该机制可能是肠道平滑肌收缩适应过程中的能量补偿通路[44]。
动力蛋白相关蛋白1(DRP1)与OPA1调控能量分布与形态稳定性:DRP1与OPA1构成线粒体动力学调节核心。肠道动力障碍模型中观察到DRP1活性增强、OPA1表达下降,造成线粒体碎片化、ATP合成能力下降,是功能性胃肠病中的常见现象[45]。
2.1.2 钙稳态失衡线粒体融合蛋白Mitofusin 1/2(MFN1/2)-MCU-NCLX通路调控线粒体钙动态:MFN1/2通过调控线粒体与内质网间Ca2+转运,维持平滑肌钙稳态。MFN1缺失会抑制MCU(线粒体钙单向转运体)表达、增强NCLX活性,导致钙输出增强,影响ATP生成与肠道平滑肌收缩功能[25, 46]。
Ca2+-IP3R耦联反馈通路调控肌质网-线粒体钙信号:线粒体通过其钙摄取能力调控肌质网IP3受体(IP3R)簇的放电活动,进而影响Ca2+信号的空间和时间特征。干扰线粒体膜电位或MCU活性会显著减弱局部钙信号,削弱肠道平滑肌细胞的收缩效应[25, 47]。
2.1.3 线粒体ROS(mtROS)信号异常mtROS-HIF-1α/NF-κB-Prohibitin2通路驱动氧化应激转录调控:mtROS在维持肠道平滑肌表型方面起双向作用。高水平mtROS可以抑制抗增殖蛋白2(PHB2)介导的促收缩蛋白表达,诱导肠道平滑肌表型去分化、功能减退。此外,mtROS还通过激活HIF-1α、NF-κB等转录因子,放大炎症反应并诱导细胞凋亡[48]。
mtROS-TOMM40-Myocardin通路调控平滑肌表型转化:除Prohibitin-2外,mtROS还通过上调线粒体跨膜转运蛋白TOMM40,协同抑制心肌素(Myocardin)及促收缩型miRNAs(如miR-145),促使平滑肌细胞从收缩型向合成型表型转变,造成功能下降和结构失稳。这在肺动脉高压及血管重构等疾病中尤为明显[49]。
mtROS-H2O2微区激活L型钙通道:研究发现,线粒体靠近于细胞膜下皮层区域时,可以局部放大H2O2信号,激活L型钙通道,增强收缩性Ca2+信号,从而影响血管平滑肌或胃肠道平滑肌张力[50]。
2.1.4 其他DRP1-AKT-细胞周期蛋白(cyclin D)通路调控线粒体动力学与细胞增殖:此通路参与病理性的平滑肌增殖。在血管或胃肠平滑肌增生相关病理中,DRP1介导的线粒体分裂促进cyclin D及Akt磷酸化激活,增强细胞增殖活性,破坏正常节律性与应激反应能力[51-52]。
一磷酸腺苷活化蛋白激酶(AMPK)-肝激酶B1(LKB1)-钙/钙调蛋白依赖性蛋白激酶激酶β(CaMKKβ)通路调节代谢适应与凋亡:AMPK是细胞内重要的能量感应器,在能量耗竭、ATP/AMP(一磷酸腺苷)比值下降或高糖等代谢应激条件下被快速磷酸化激活,其下游可以通过LKB1和CaMKKβ等激酶通路共同调控能量代谢、脂肪酸氧化、葡萄糖摄取以及线粒体生物合成,并进一步参与自噬、细胞周期阻滞与程序性细胞死亡等过程,从而在应激环境中协调细胞的代谢适应与存活或凋亡的命运决策。糖尿病胃轻瘫模型显示,AMPK活化可以促进线粒体代谢障碍与糖酵解增强,并诱导肠道平滑肌细胞凋亡,降低肠蠕动[53-54]。
线粒体微小RNA(Mito-miR)信号介导线粒体-核反馈与表观遗传调控:Mito-miR信号与线粒体-核反馈通路中的表观遗传调控息息相关。线粒体通过合成或调节mito-miRs(如miR-145、miR-21、miR-378、miR-761)参与表型转化与代谢调控,抑制线粒体分裂可以显著下调促增殖miR-21表达,同时恢复miR-145水平,促进收缩型表型维持[55-56]。
2.2 肠道平滑肌功能失调对线粒体功能的反向反馈机制除线粒体功能障碍可以作为肠道平滑肌动力减弱的重要起始因素外,越来越多研究表明,肠道平滑肌本身的功能失调亦可以通过多种途径反向作用于线粒体,诱发或加重其功能障碍。在肠道平滑肌长期处于收缩节律紊乱、张力异常、钙稳态失衡或炎症激活等病理状态时,可以通过钙信号过载、线粒体氧化应激增强、动力学重构及代谢重编程等机制,持续损害线粒体结构与能量代谢功能。这一过程使线粒体由功能受损的细胞器进一步转变为放大病理信号的关键节点,从而与肠道平滑肌功能障碍之间形成相互促进、不断强化的病理性正反馈环路。
2.2.1 肠道平滑肌功能失调诱导钙超载在肠道平滑肌张力增高或电活动失衡条件下,Ca2+内流增加引起胞质钙超载,过量Ca2+被线粒体摄取后诱导线粒体通透性转换孔(mPTP)开放,造成线粒体膜电位丧失和ROS激增,进一步损伤线粒体呼吸链[40]。在模拟微重力导致的平滑肌过度收缩模型中发现,肌张力增强与Ca2+负载上升伴随线粒体融合蛋白MFN1/2表达下降、DRP1上调,提示钙负荷对线粒体形态调控具有双向影响。此外,线粒体膜电位降低、钙/磷酸盐积聚会协同激活ROS生成并损伤呼吸链复合物Ⅰ功能,减少ATP合成[57]。
2.2.2 肠道平滑肌功能失调抑制线粒体自噬当平滑肌长期处于高张力、代谢负荷或凋亡刺激下,线粒体自噬机制会受到抑制,致使损伤的线粒体积聚,引发细胞炎症与代谢重编程。在肌肉张力失衡模型中发现,线粒体融合蛋白OPA1和MFN1/2表达下降,DRP1介导的线粒体碎片化增强,进一步诱导线粒体网络分裂、功能丧失。而过度分裂后的线粒体难以通过PINK1/Parkin途径被有效清除,导致功能障碍的线粒体积聚,引发代谢毒性累积和进一步的肠道平滑肌损伤[58]。
2.2.3 肠道平滑肌功能失调驱动线粒体代谢重编程在肌肉疾病或慢性功能低下时,肠道平滑肌细胞功能失调会持续进展,线粒体代谢程序被迫发生重新配置,由OXPHOS逐步转向高度依赖糖酵解的能量模式。在这一过程中,线粒体呼吸链复合物Ⅰ和Ⅲ的功能障碍是首要触发因素,造成电子传递效率下降、ATP生成能力受限及代偿性乳酸代谢增强,继而引发细胞内pH下降和ROS累积。该代谢重编程不仅影响能量供需,而且通过激活HIF-1α等应激转录因子诱导肠道平滑肌细胞向合成型、增殖型,甚至炎症型表型转化,进一步抑制其收缩功能[59-60]。
线粒体分裂蛋白DRP1在此过程中表现出显著上调,其过度激活不仅促进线粒体裂解和功能碎片化,还抑制OPA1介导的嵴结构维持与mtDNA稳定,从而削弱线粒体的OXPHOS潜力。同时,线粒体裂解伴随的mtROS升高亦通过信号放大效应激活c-Myc、NF-κB等代谢转录因子,加剧糖酵解酶(如HK2、PFKFB3)表达升高,最终促使代谢重编程进入稳定维持期[61-62]。
由此可见,肠道平滑肌的功能失调不仅表现为收缩失能,更可通过代谢通路的重构与线粒体网络动态变化,引发持续性的细胞代谢模式和表型改变,形成恶性循环。
3 线粒体功能障碍与阳虚型便秘的关系 3.1 阳气与线粒体的内在关联根据中医基础理论,“阳气”是人体之气中具有温煦、推动与兴奋作用的部分,是维持生命活动的动力之源[63]。而现代生命科学中,细胞功能的基本能量来源正是线粒体,它是维持机体体温、运动、代谢、免疫等系统正常运行的关键结构。从功能角度看,两者在维持基础体温、调控昼夜节律、参与免疫调节、影响衰老进程等方面的作用高度相似。
“阳虚”(亦称“阳气虚”)则特指阳气功能减退、导致脏腑功能衰退、机体呈现虚寒状态的病理证候[64]。在临床辨证中,“阳虚”与“阳气虚”常可互用。在此状态下的畏寒怕冷、易疲劳、四肢不温等症状,也常见于线粒体功能障碍相关疾病,如慢性疲劳综合征、老年性代谢减退等。
在1项针对寒证与线粒体代谢的观察性临床研究中发现,阳虚与寒证体质人群的线粒体功能显著低下,包括氧消耗率(OCR)、ATP含量、线粒体酶活性等均显著低于平和体质人群[63]。另外1项慢性疲劳综合征为模型的研究发现,阳虚患者体内ATP生成速率、线粒体内膜流动性显著下降,而应用杜仲、淫羊藿、补骨脂等“阳药”被证实能够通过促进线粒体膜流动性、提高ATP合成速率来强化能量代谢,人参、黄芪等则更多通过提升谷胱甘肽水平、抗氧化保护线粒体结构[65]。在阳虚动物模型中,其肾上腺皮质、胃肠黏膜、骨骼肌等组织中线粒体数量显著减少,形态异常(如嵴断裂、肿胀)、膜电位降低、ATP合成能力下降。使用温阳的附子类方剂干预后,可以显著改善上述线粒体形态与功能异常,进一步印证“阳虚状态与线粒体功能障碍存在高度相关性”的观点[64-65]。
需要指出的是,线粒体功能障碍并非对各组织器官产生均等影响。相较于结构稳定、能量消耗相对恒定的组织,对持续能量供应和钙信号高度依赖的平滑肌系统更易受到线粒体功能下降的影响。肠道平滑肌作为维持胃肠推进和节律性蠕动的关键执行单元,其收缩活动高度依赖ATP持续供给及线粒体介导的钙稳态调控。因此,在“阳虚/线粒体功能低下”的背景下,肠道平滑肌功能障碍可能成为阳虚状态在胃肠系统中的优先表现形式之一。
3.2 阳虚型便秘与肠道平滑肌动力障碍如前文所述,基于上述阳气与线粒体在能量代谢上的共性,可以进一步推论:当机体处于阳虚状态时,其伴发的线粒体功能障碍若累及肠道平滑肌细胞,将直接导致该细胞能量供应不足、钙稳态与氧化还原平衡失调,从而引发肠道平滑肌收缩无力与节律紊乱,这即是阳虚型便秘中“肠道动力障碍”的现代病理学实质。
本文所研究的“阳虚型便秘”,即指以脾肾阳气亏虚、温煦推动无力为核心病机的便秘证型。阳虚型便秘作为一种常见的便秘类型,通常表现为大便艰涩不易排出,便后无力,腹部胀满,腹痛喜温喜按,四肢不温,或腰膝酸冷,小便清长,面色白,舌淡苔白,脉沉迟[66]。这一系列症状均指向机体基础能量代谢与动力供应的衰退。从功能角度看,此种以“能量代谢动力不足”为特征的“阳虚”状态,与西医中线粒体功能障碍所导致的细胞及组织层面“低能耗-低功能”表现,在病理生理层面高度契合。在传统中医理论中,虽然未明确提及“肠道平滑肌功能障碍”这一西医概念,但诸多相关症状可以归属于“脾阳虚”“肾阳虚”等“阳虚”证型范畴。中医认为脾主运化,脾胃为后天之本、气血生化之源。而肾为先天之本,脾胃的运化功能依赖于肾阳的温养[67]。若肾阳虚,不能温煦脾胃,导致脾阳虚弱,运化失职,则可出现腹胀、腹痛、便秘或泄泻等肠道传导功能异常的表现,这些症状与肠道平滑肌功能障碍所引发的临床表现高度相似。随着现代生活节奏加快,人们饮食结构发生改变,过食生冷寒凉之物极为普遍。从中医角度看,生冷寒凉之品易损伤脾胃阳气,导致脾阳虚衰,运化无权,使得肠道平滑肌功能障碍的发生率逐渐上升[68]。在动物实验中,通过“饮食不节+过劳+泻下”三联刺激复制的脾阳虚模型表现出胃排空延迟、肠推进率下降、肠道平滑肌收缩力减弱等一系列胃肠动力功能障碍表现[69]。
此外,现代流行病学调查显示,阳虚型在肠道动力障碍患者中占比达到50%左右,与年龄增长呈显著正相关[9]。这一数据提示,阳虚型便秘是慢性便秘中的一个主要证型。虽然阴虚、血虚等其他证型也可能导致便秘,但其病理机制可能侧重于“津枯肠燥”“濡润不足”等不同方面。本研究聚焦于阳虚型便秘,正是因为其“畏寒、乏力、能量代谢动力不足”的核心表现与病机,和线粒体功能障碍导致的“低能耗-低功能”状态在理论上的关联最为直接和典型。这也为日后将研究聚焦于该证型,并深入探究其生物分子学机制提供了依据。
3.3 温阳疗法改善肠道平滑肌动力障碍、治疗阳虚型便秘从病理生理层面看,阳虚型便秘所表现出的排便无力、肠道传导减慢及蠕动节律紊乱,其直接执行基础在于肠道平滑肌收缩功能减弱。因此,能够改善肠道推进率、收缩幅度与节律的干预措施,实质上均作用于肠道平滑肌这一核心目标。
研究表明,采用温补脾阳法的中药方剂,如附子理中丸、香砂六君子汤、四君子汤等治疗脾虚阳虚型功能性消化不良患者,可以显著减轻患者胃脘胀满、疼痛、嗳气、纳差等症状,改善胃排空延迟、小肠推进率下降及结肠传输时间延长等表现,并同步提高胃肠收缩频率与协调性,且治疗后的中医症状积分明显降低,疗效优于常规西药治疗组[70]。另有研究发现,针刺足三里、中脘等穴位配合温和灸神阙穴,能够改善脾肾阳虚型便秘患者肠道易激惹症状,调节胃肠动力,缩短结肠传输时间,提高临床疗效[71-72]。这些结果均提示,温阳疗法能够有效恢复肠道平滑肌的动力输出能力,从而缓解阳虚型便秘相关的动力障碍症状。上述干预方式虽然作用形式不同,但其共同指向均为肠道平滑肌收缩功能的恢复。值得注意的是,肠道平滑肌动力学改善并非单独发生,而是与肠道平滑肌细胞能量代谢、钙稳态及氧化应激状态改善密切相关,这为进一步从线粒体层面解析温阳疗法的作用机制提供了重要线索。
从机制层面看,上述温阳疗法在改善肠道平滑肌收缩功能的同时,亦可能通过纠正平滑肌异常张力与钙信号失衡,间接阻断其对线粒体功能的反向损伤过程,从而在“线粒体功能障碍-平滑肌动力失调”的双向作用中发挥整体调节效应。
3.4 温阳疗法改善线粒体功能的机制研究如前文所述,阳虚型便秘的病机与肠道平滑肌线粒体功能障碍机制在能量匮乏、钙稳态失调和氧化应激等方面存在共性。因此,温阳疗法改善阳虚型便秘的现代生物学机制,其关键切入点之一在于修复肠道平滑肌的线粒体功能。以下研究从不同侧面证实,温阳中药及其活性成分正是通过靶向调控线粒体的能量生成、钙稳态与氧化还原平衡3个核心功能,从而恢复肠道平滑肌动力,为“阳虚型便秘-肠道平滑肌线粒体功能障碍”这一假说提供了直接的实验支持。
3.4.1 中药活性成分改善线粒体功能机制研究为清晰概述温阳中药活性成分靶向线粒体的核心机制,现将主要证据归纳见开放科学(资源服务)标识码(OSID)。
3.4.1.1 提升能量生成,针对阳虚之“动力不足”能量代谢不足是线粒体功能障碍导致肠道平滑肌收缩乏力的核心机制之一(见“2.1.1”项)。多项研究表明,温阳及益气类中药可以通过增强线粒体ATP生成能力,为肠道平滑肌持续、节律性收缩提供必要的能量基础。针对阳虚型便秘核心的“能量生成不足”,多种温阳成分靶向增强线粒体ATP合成。如附子(乌头碱)与肉桂(肉桂醛)通过调控AMPK/PGC-1信号通路,促进线粒体生物合成与呼吸链功能,从根源提升ATP产量[73-75]。这一过程依赖于沉默信息调节因子1(SIRT1)介导的线粒体生物合成激活,通过去乙酰化修饰增强核编码线粒体基因(NRF1和TFAM)等的表达[75]。另有研究证明,附子-肉桂配伍可以通过激活AMPK的经典下游通路(cAMP/PKA通路)显著提升ATP合成与肠道推进力[76]。黄芪具有升阳补气功效,其重要成分黄芪甲苷Ⅳ也能够通过激活AMPK-PGC-1α通路,上调NRF1表达,促进线粒体生物合成,恢复细胞ATP含量[77]。人参皂苷Rg1可以通过激活SIRT1/PGC-1α轴增强线粒体OXPHOS,显著提升ATP生成与抗氧化能力,改善能量供应不足状态[78]。淫羊藿苷通过上调AMPK表达促进线粒体DNA复制与呼吸链蛋白合成,提升线粒体能量代谢效率,改善能量缺乏引起的细胞功能低下[79]。此外,温阳药物对解偶联蛋白2(UCP2)的调节作用可以通过减少线粒体质子泄漏来优化线粒体能量转化效率这一作用机制也得到验证[77, 80-81]。
在肠道平滑肌细胞高度依赖ATP驱动收缩与离子转运的背景下,上述研究从能量代谢层面提示,温阳类中药可能通过恢复线粒体ATP供给,直接改善平滑肌动力不足这一阳虚型便秘的重要病理环节。
3.4.1.2 调节钙稳态,改善兴奋-收缩耦联线粒体钙稳态失衡是导致平滑肌兴奋-收缩耦联障碍的关键因素之一(见“2.1.2”项)。相关研究表明,多种温阳中药活性成分可以通过调控线粒体钙摄取与释放,减轻钙超载对线粒体和细胞功能的损害。甘草具有补中益气功效,其主要三萜类成分18β-甘草次酸(18β-GA)以及其母体成分甘草酸(glycyrrhizin),已经被证明在平滑肌细胞内Ca2+过载模型中能降低游离Ca2+含量,减轻细胞内钙超载[82]。而甘草中另一类经典活性成分黄酮类甘草异黄酮光甘草定(glabridin)也被证明能够调节Ca2+浓度变化,验证了其对线粒体的保护作用[83]。陈皮性温,可以理中健脾,它的主要黄酮类成分橙皮苷能够通过抑制内质网(ER)与线粒体接触位点MAMs上的IP3R/GRP75/VDAC1-MCU钙输入轴,降低线粒体钙过载并改善肠道线粒体功能[84]。柚皮苷通过减少线粒体对Ca2+的膜电位依赖性积累,抑制Ca2+诱导的线粒体通透性转变孔MPTP开放,阻止膜去极化与呼吸耦联破坏,实现线粒体功能保护[85]。
该类研究提示,通过缓解线粒体钙稳态紊乱,温阳中药可能有助于恢复肠道平滑肌正常的兴奋-收缩耦联过程,从而改善动力障碍状态。
3.4.1.3 减轻氧化应激与抑制凋亡,保护平滑肌结构与功能线粒体ROS过度积聚及凋亡信号激活是导致肠道平滑肌功能减退和结构破坏的重要病理过程(见“2.1.3”与“2.1.4”项)。肉桂中的肉桂醛可以在缺血/缺氧等氧化应激模型中提高抗氧化酶,如SOD、GPx等的活性并降低ROS与脂质过氧化产物水平,减轻氧化损伤[86]。五味子性温,其主要生物活性成分为二苯并环辛烯类木脂素,它们对细胞具有直接/间接抗氧化作用,其中五味子甲素通过清除ROS、保护线粒体功能来维持能量代谢与细胞存活[87]。五味子乙素可以显著降低ROS、增强谷胱甘肽相关抗氧化能力,并维持线粒体完整性[88]。当归中的活性成分阿魏酸(FA)能够显著降低ROS水平,不仅直接清除自由基,还能够提高SOD、CAT、GPx等抗氧化酶活性,增强内源性抗氧化防御,进而恢复线粒体膜电位,抑制凋亡,稳定线粒体功能并有助于维持ATP与能量代谢稳态[89-90]。黄芪甲苷可以通过诱导p62磷酸化、促进Nrf2核转位及其下游抗氧化基因表达,降低线粒体ROS水平并改善线粒体功能[91]。人参皂苷Rg1已经被验证通过多靶点降低线粒体ROS并保护线粒体:一方面下调miR-144,促进Nrf2核转位并激活ARE抗氧化基因,上调HO-1/NQO1、SOD、GPx等酶以抑制ROS产生[92-93];另一方面,作为SIRT1激活剂增强PINK1/Parkin介导的线粒体自噬,清除受损线粒体以减少线粒体ROS源头[94],同时维持线粒体嵴结构和呼吸链效率,从而整体改善线粒体功能与细胞耐受氧化应激能力[95]。肉苁蓉可以温补肾阳,其标志性成分毛蕊花糖苷(ACT)可以在氧化应激/损伤模型中降低线粒体ROS,恢复线粒体膜电位,并通过抑制线粒体介导的Caspase-9/3凋亡通路提高细胞存活率[96];ACT还通过抑制脯氨酸羟化酶2(PHD2)上调BNIP3-PINK1/Parkin等线粒体自噬相关蛋白,促进线粒体自噬清除受损线粒体[97]。巴戟天温肾阳、强筋骨,其重要成分巴戟天寡糖(MOOs)已被证实可以增强线粒体SOD2活性,促进GPX4向线粒体转位,抑制NOX4降低线粒体氧化压力,抑制线粒体相关铁死亡[98];还可以上调Mfn2,激活线粒体自噬,从而清除受损线粒体[99]。干姜中的6-姜酚在多种损伤/应激模型中可以恢复线粒体膜电位并减轻线粒体肿胀和膜破裂,维持线粒体结构与功能[100];同时通过上调Bcl-2、下调Bax及降低Caspase-3/cleaved-Caspase-3水平,抑制线粒体依赖性凋亡,从而实现对“线粒体-凋亡轴”的修复与细胞保护[101]。肉桂提取物桂皮醛在细胞氧化或毒性损伤中主要通过两条线粒体相关路径发挥抗凋亡作用:其一是直接稳定线粒体功能,即维持线粒体膜电位,抑制过量ROS累积,从而阻止线粒体外膜通透性异常升高,减少细胞色素C从线粒体释放到胞质,进而抑制Caspase-3的裂解/激活,最终降低线粒体内源性凋亡通路的启动[102];其二是通过线粒体质量控制减少线粒体应激信号的产生:桂皮醛可以促进PINK1/Parkin相关的线粒体自噬,清除受损、持续产生ROS的线粒体亚群,使线粒体群体保持更高的结构与功能完整性[103-104]。
在肠道平滑肌长期处于低能量与氧化应激状态的阳虚型便秘背景下,这些作用机制为温阳疗法通过“保护线粒体-维持平滑肌结构与功能”改善肠道动力提供了更多科学依据。
3.4.2 中药复方改善线粒体功能的机制研究相较单一成分,中药复方具有多通路协同调控优势,其对线粒体功能的整体改善效应,可能更契合阳虚型便秘中复杂、系统的病理特征。为清晰概述温阳类复方靶向线粒体的核心机制,现将主要证据归纳,见OSID。
结合肠道平滑肌高度依赖线粒体功能维持收缩与节律的生理特点,研究从复方层面进一步支持了“通过调控肠道平滑肌线粒体功能改善阳虚型便秘”的核心假设。
综上所述,阳虚不仅在中医辨证体系中与胃肠道疾病密切相关,其现代病理基础亦可归结为线粒体功能障碍对肠道平滑肌动力存在重要影响。无论是单体成分还是复方,温阳中药的现代药理作用均高度聚焦于纠正线粒体能量代谢障碍、稳定钙信号、抑制氧化损伤与阳虚型便秘病理生理紧密关联的核心环节,进而改善肠道平滑肌的收缩与节律功能,这些证据不再是孤立的药理发现,而是为“肠道平滑肌线粒体-肠道平滑肌功能障碍-阳虚型便秘”的发病机制提供了实验证据,强有力地支持了“肠道平滑肌线粒体功能障碍是阳虚型便秘的关键微观病理基础,而温阳疗法的起效机制在于修复此靶点”这一核心假设。
4 总结与展望随着西医对阳虚型便秘机制认识的不断深入,线粒体作为能量代谢与细胞信号调控核心的角色日益突显。作为肠道平滑肌细胞代谢、信号和结构稳态的核心枢纽,线粒体的功能状态直接决定着平滑肌的收缩能力、节律性蠕动以及细胞存活与分化状态。本研究系统梳理了线粒体功能障碍通过能量失衡、钙信号紊乱、ROS积聚、凋亡激活及动力学改变等机制与肠道平滑肌功能障碍的关系,提出“肠道平滑肌线粒体-肠道平滑肌功能障碍-阳虚型便秘”这一视角,为中医辨证体系与现代分子机制的融合搭建了可以验证的桥梁。
然而,目前研究多集中于动物模型或初步机制探索,临床证据仍显薄弱,机制链条的完整性尚需进一步证实。未来应从以下几个方面展开深入研究:其一,构建更符合中医证型特征的线粒体功能障碍动物模型,提升研究的一致性;其二,结合代谢组学、线粒体组学等技术揭示“肠道平滑肌线粒体-肠道平滑肌功能障碍-阳虚型便秘”之间的多维关联;其三,加强临床与基础研究的双向转化,探索中药复方与肠道平滑肌线粒体及肠道平滑肌信号通路和靶点之间的因果联系,推动个体化干预策略的形成;其四,利用类器官、肠道平滑肌细胞特异性线粒体调控模型等新技术,更精确地模拟和验证“阳虚”病理状态下肠道平滑肌线粒体的功能变化及中药的干预节点。
综上所述,“阳虚”不仅是中医辨证的核心概念,也可能在线粒体功能受损所引起的肠道平滑肌功能障碍中扮演重要角色。挖掘其生物学基础,不仅有助于推动中医理论现代化,也为阳虚型便秘等慢性病的治疗开辟了更具整合性与前瞻性的路径。
| [1] |
BASSOTTI G, USAI SATTA P, BELLINI M. Chronic idiopathic constipation in adults: A review on current guidelines and emerging treatment options[J]. Clinical and Experimental Gastroenterology, 2021, 14: 413-428. DOI:10.2147/CEG.S256364 |
| [2] |
BHARUCHA A E, LACY B E. Mechanisms, evaluation, and management of chronic constipation[J]. Gastroenterology, 2020, 158(5): 1232-1249. DOI:10.1053/j.gastro.2019.12.034 |
| [3] |
ZHANG T, ZULLO A R, JAMES H O, et al. The burden and treatment of chronic constipation among US nursing home residents[J]. Journal of the American Medical Directors Association, 2023, 24(8): 1247-1252. DOI:10.1016/j.jamda.2023.05.006 |
| [4] |
BARBERIO B, JUDGE C, SAVARINO E V, et al. Global prevalence of functional constipation according to the Rome criteria: A systematic review and meta-analysis[J]. The Lancet Gastroenterology & Hepatology, 2021, 6(8): 638-648. |
| [5] |
SONG B G, LEE Y C, MIN Y W, et al. Risk of domperidone induced severe ventricular arrhythmia[J]. Scientific Reports, 2020, 10(1): 12158. DOI:10.1038/s41598-020-69053-4 |
| [6] |
BOR S, LEHERT P, CHALBAUD A, et al. Efficacy of pinaverium bromide in the treatment of irritable bowel syndrome: A systematic review and meta-analysis[J]. Therapeutic Advances in Gastroenterology, 2021, 14: 108-116. |
| [7] |
WANG L, WU F, HONG Y, et al. Research progress in the treatment of slow transit constipation by traditional Chinese medicine[J]. Journal of Ethnopharmacology, 2022, 290: 1150-1175. |
| [8] |
中华中医药学会脾胃病分会. 慢性便秘中医诊疗共识意见[J]. 北京中医药, 2011, 30(1): 3-7. |
| [9] |
杨勇, 丁曙晴, 杨光, 等. 功能性便秘中医证候与症状严重度的量化研究[J]. 南京中医药大学学报, 2015, 31(1): 24-27. |
| [10] |
ZHAO Q, CHEN Y, XU D, et al. Action mode of gut motility, fluid and electrolyte transport in chronic constipation[J]. Frontiers in Pharmacology, 2021, 12: 630249. DOI:10.3389/fphar.2021.630249 |
| [11] |
GILBERT M A, SCHULTZ-ROGERS L, RAJAGOPALAN R, et al. Protein-elongating mutations in MYH11 are implicated in a dominantly inherited smooth muscle dysmotility syndrome with severe esophageal, gastric, and intestinal disease[J]. Human Mutation, 2020, 41(5): 973-982. DOI:10.1002/humu.23986 |
| [12] |
罗鹏. 大承气汤对多器官功能障碍综合征大鼠小肠平滑肌线粒体凋亡途径和NF-κB、COX-2蛋白表达影响[D]. 大连: 大连医科大学, 2014.
|
| [13] |
LIU X, YANG K, JIA Y, et al. Chaihushugan powder regulates the gut microbiota to alleviate mitochondrial oxidative stress in the gastric tissues of rats with functional dyspepsia[J]. Frontiers in Immunology, 2025, 16: 1549554. DOI:10.3389/fimmu.2025.1549554 |
| [14] |
张天华, 赵莎彤, 黎晓宇, 等. 基于PI3K/Akt/mTOR信号通路探讨电针对糖尿病胃轻瘫大鼠胃窦部Cajal间质细胞自噬的影响[J]. 针刺研究, 2022, 47(12): 1060-1067. |
| [15] |
ANAND R, REICHERT A S, KONDADI A K. Emerging roles of the MICOS complex in cristae dynamics and biogenesis[J]. Biology, 2021, 10(7): 600. DOI:10.3390/biology10070600 |
| [16] |
NAKANO K, YAMAMOTO M, YAMADA Y, et al. Mitochondrial structural abnormalities and cardiac reverse remodeling in patients with systolic dysfunction[J]. Circulation Journal, 2024, 89(1): 101-108. DOI:10.1253/circj.CJ-24-0451 |
| [17] |
ASUNCIÓN-ALVAREZ D, PALACIOS J, YBAÑEZ-JULCA R O, et al. Calcium signaling in endothelial and vascular smooth muscle cells: Sex differences and the influence of estrogens and androgens[J]. American Journal of Physiology: Heart and Circulatory Physiology, 2024, 326(4): H950-H970. DOI:10.1152/ajpheart.00600.2023 |
| [18] |
CLEMENTE-SUÁREZ V J, REDONDO-FLÓREZ L, BELTRÁN-VELASCO A I, et al. Mitochondria and brain disease: A comprehensive review of pathological mechanisms and therapeutic opportunities[J]. Biomedicines, 2023, 11(9): 2488. DOI:10.3390/biomedicines11092488 |
| [19] |
ZHAO R, JIANG S, ZHANG L, et al. Mitochondrial electron transport chain, ROS generation and uncoupling (review)[J]. International Journal of Molecular Medicine, 2019, 44(1): 3-15. |
| [20] |
JAVADOV S, JANG S, CHAPA-DUBOCQ X R, et al. Mitochondrial respiratory supercomplexes in mammalian cells: Structural versus functional role[J]. Journal of Molecular Medicine, 2021, 99(1): 57-73. DOI:10.1007/s00109-020-02004-8 |
| [21] |
ARDALAN A, SMITH M D, JELOKHANI-NIARAKI M. Uncoupling proteins and regulated proton leak in mitochondria[J]. International Journal of Molecular Sciences, 2022, 23(3): 1528. DOI:10.3390/ijms23031528 |
| [22] |
TAM B T, MURPHY J, KHOR N, et al. Acetyl-CoA regulation, OXPHOS integrity and leptin levels are different in females with childhood vs adulthood onset of obesity[J]. Endocrinology, 2020, 161(11): bqaa142. DOI:10.1210/endocr/bqaa142 |
| [23] |
RASSIER D E, MÅNSSON A. Mechanisms of myosin Ⅱ force generation: Insights from novel experimental techniques and approaches[J]. Physiological Reviews, 2025, 105(1): 1-93. DOI:10.1152/physrev.00014.2023 |
| [24] |
HULSURKAR M M, LAHIRI S K, KARCH J, et al. Targeting calcium-mediated inter-organellar crosstalk in cardiac diseases[J]. Expert Opinion on Therapeutic Targets, 2022, 26(4): 303-317. DOI:10.1080/14728222.2022.2067479 |
| [25] |
BOYMAN L, GREISER M, LEDERER W J. Calcium influx through the mitochondrial calcium uniporter holocomplex, MCU(cx)[J]. Journal of Molecular and Cellular Cardiology, 2021, 151: 145-154. DOI:10.1016/j.yjmcc.2020.10.015 |
| [26] |
BOCZEK T, SOBOLCZYK M, MACKIEWICZ J, et al. Crosstalk among calcium ATPases: PMCA, SERCA and SPCA in mental diseases[J]. International Journal of Molecular Sciences, 2021, 22(6): 2785. DOI:10.3390/ijms22062785 |
| [27] |
WALKON L L, STRUBBE-RIVERA J O, BAZIL J N. Calcium overload and mitochondrial metabolism[J]. Biomolecules, 2022, 12(12): 1891. DOI:10.3390/biom12121891 |
| [28] |
GAETANI S, GALZIGNATI L, MARCATI M, et al. Mitochondrial function as related to psychological distress in health care professionals[J]. Psychosomatic Medicine, 2022, 84(1): 40-49. DOI:10.1097/PSY.0000000000001000 |
| [29] |
PATTON B L, ZHU P, ELSHEIKH A, et al. Dynamic duo: Kir6 and SUR in K(ATP) channel structure and function[J]. Channels, 2024, 18(1): 2327708. DOI:10.1080/19336950.2024.2327708 |
| [30] |
AL KURY L T. Modulatory effect of medicinal plants and their active constituents on ATP-sensitive potassium channels(K(ATP)) in diabetes[J]. Pharmaceuticals, 2023, 16(4): 523. DOI:10.3390/ph16040523 |
| [31] |
O'FLAHERTY C, SCARLATA E. Oxidative stress and reproductive function: The protection of mammalian spermatozoa against oxidative stress[J]. Reproduction, 2022, 164(6): F67-F78. DOI:10.1530/REP-22-0200 |
| [32] |
NAM Y, KIM H J, KWON O. Acute and prolonged effects of Bacillus amyloliquefaciens GF424-derived SOD on anti-oxidant defense in healthy individuals challenged with intense aerobic exercise[J]. Free Radical Biology and Medicine, 2024, 224: 484-493. DOI:10.1016/j.freeradbiomed.2024.09.015 |
| [33] |
SALDMANN F, VILTARD M, LEROY C, et al. The naked mole rat: A unique example of positive oxidative stress[J]. Oxidative Medicine and Cellular Longevity, 2019, 2019: 4502819. |
| [34] |
CATTERALL W A, LENAEUS M J, GAMAL EL-DIN T M. Structure and pharmacology of voltage-gated sodium and calcium channels[J]. Annual Review of Pharmacology and Toxicology, 2020, 60: 133-154. DOI:10.1146/annurev-pharmtox-010818-021757 |
| [35] |
MARCUCCI L. Muscle mechanics and thick filament activation: An emerging two-way interaction for the vertebrate striated muscle fine regulation[J]. International Journal of Molecular Sciences, 2023, 24(7): 6265. DOI:10.3390/ijms24076265 |
| [36] |
ZHAO Y, PENG C, LAI R, et al. The SNPs of mitochondrial DNA displacement loop region and mitochondrial DNA copy number associated with risk of polymyositis and dermatomyositis[J]. Scientific Reports, 2022, 12(1): 5903. DOI:10.1038/s41598-022-09943-x |
| [37] |
WEI F, NIAN Q, ZHAO M, et al. Natural products and mitochondrial allies in colorectal cancer therapy[J]. Biomedicine & Pharmacotherapy, 2023, 167: 115473. |
| [38] |
SEKAR G, OJOAWO A, MOLDOVEANU T. Protein-protein and protein-lipid interactions of pore-forming BCL-2 family proteins in apoptosis initiation[J]. Biochemical Society Transactions, 2022, 50(3): 1091-1103. DOI:10.1042/BST20220323 |
| [39] |
CZABOTAR P E, GARCIA-SAEZ A J. Mechanisms of BCL-2 family proteins in mitochondrial apoptosis[J]. Nature Reviews Molecular Cell Biology, 2023, 24(10): 732-748. DOI:10.1038/s41580-023-00629-4 |
| [40] |
CUI Y, PAN M, MA J, et al. Recent progress in the use of mitochondrial membrane permeability transition pore in mitochondrial dysfunction-related disease therapies[J]. Molecular and Cellular Biochemistry, 2021, 476(1): 493-506. DOI:10.1007/s11010-020-03926-0 |
| [41] |
VITALE I, PIETROCOLA F, GUILBAUD E, et al. Apoptotic cell death in disease-current understanding of the NCCD 2023[J]. Cell Death and Differentiation, 2023, 30(5): 1097-1154. |
| [42] |
PARSEH S, SHAKERIAN S, REZA TABANDEH M, et al. An 8-week study on the effects of high and moderate-intensity interval exercises on mitochondrial MOTS-C changes and their relation to metabolic markers in male diabetic sand rats[J]. Diabetes Research and Clinical Practice, 2024, 212: 111656. DOI:10.1016/j.diabres.2024.111656 |
| [43] |
LEWIS M T, KASPER J D, BAZIL J N, et al. Skeletal muscle energetics are compromised only during high-intensity contractions in the Goto-Kakizaki rat model of type 2 diabetes[J]. American Journal of Physiology: Regulatory, Integrative and Comparative Physiology, 2019, 317(2): R356-R368. DOI:10.1152/ajpregu.00127.2019 |
| [44] |
ATLANTE A, VALENTI D. A walk in the memory, from the first functional approach up to its regulatory role of mitochondrial bioenergetic flow in health and disease: Focus on the adenine nucleotide translocator[J]. International Journal of Molecular Sciences, 2021, 22(8): 4164. DOI:10.3390/ijms22084164 |
| [45] |
BAO L, YU Y, GONZÁLEZ-ACERA M, et al. Epithelial OPA1 links mitochondrial fusion to inflammatory bowel disease[J]. Science Translational Medicine, 2025, 17(781): n8699. DOI:10.1126/scitranslmed.adn8699 |
| [46] |
LU X, WANG Y, GENG N, et al. Dysregulated mitochondrial calcium causes spiral artery remodeling failure in preeclampsia[J]. Hypertension, 2024, 81(11): 2368-2382. DOI:10.1161/HYPERTENSIONAHA.124.23046 |
| [47] |
GARBINCIUS J F, LUONGO T S, ELROD J W. The debate continues-what is the role of MCU and mitochondrial calcium uptake in the heart?[J]. Journal of Molecular and Cellular Cardiology, 2020, 143: 163-174. DOI:10.1016/j.yjmcc.2020.04.029 |
| [48] |
ZHAO C, SHEN J, LU Y, et al. Dedifferentiation of vascular smooth muscle cells upon vessel injury[J]. International Immunopharmacology, 2025, 144: 113691. DOI:10.1016/j.intimp.2024.113691 |
| [49] |
PEARCE W J. Mitochondrial influences on smooth muscle phenotype[J]. American Journal of Physiology: Cell Physiology, 2024, 326(2): C442-C448. DOI:10.1152/ajpcell.00354.2023 |
| [50] |
MUNTEANU C, POPESCU C, VLĂDULESCU-TRANDAFIR A, et al. Signaling paradigms of H(2)S-induced vasodilation: A comprehensive review[J]. Antioxidants, 2024, 13(10): 1158. DOI:10.3390/antiox13101158 |
| [51] |
FIGUEROA-TOLEDO A M, GUTIÉRREZ-PINO J, CARRIEL-NESVARA A, et al. BMAL1 and CLOCK proteins exhibit differential association with mitochondrial dynamics, protein synthesis pathways and muscle strength in human muscle[J]. The Journal of Physiology, 2024, 602(23): 6403-6415. DOI:10.1113/JP285955 |
| [52] |
MAO X, GU Y, SUI X, et al. Phosphorylation of dynaminrelated protein 1(DRP1) regulates mitochondrial dynamics and skeletal muscle wasting in cancer cachexia[J]. Frontiers in Cell and Developmental Biology, 2021, 9: 673618. |
| [53] |
ZHANG M, FANG X, GUO J, et al. Effects of AMPK on apoptosis and energy metabolism of gastric smooth muscle cells in rats with diabetic gastroparesis[J]. Cell Biochemistry and Biophysics, 2019, 77(2): 165-177. DOI:10.1007/s12013-019-00870-9 |
| [54] |
CUI Y X, CHEN J H, ZHANG Z, et al. The role of AMPK in macrophage metabolism, function and polarisation[J]. Journal of Translational Medicine, 2023, 21: 892. |
| [55] |
ROSOLEN D, NUNES-SOUZA E, MARCHI R, et al. MiR-NAs action and impact on mitochondria function, metabolic reprogramming and chemoresistance of cancer cells: A systematic review[J]. Biomedicines, 2023, 11(3): 693. |
| [56] |
PUTRA A D, ANDRIJONO, WINARTO H, et al. miR-21 and miR-145 as prognostic biomarkers for radiotherapy responses in cervical cancer patients: A preliminary study[J]. International Journal of Molecular Sciences, 2024, 25(19): 10545. |
| [57] |
LIU Y, CAO X, ZHOU Q, et al. Mechanisms and counter-measures for muscle atrophy in microgravity[J]. Cells, 2024, 13(24): 2120. |
| [58] |
DASGUPTA D, DELMOTTE P, SIECK G C. Inflammation-induced protein unfolding in airway smooth muscle triggers a homeostatic response in mitochondria[J]. International Journal of Molecular Sciences, 2020, 22(1): 363. |
| [59] |
JIA Y, WANG M, MAO C, et al. COMP-prohibitin 2 interaction maintains mitochondrial homeostasis and controls smooth muscle cell identity[J]. Cell Death & Disease, 2018, 9(6): 676. |
| [60] |
LI H, ZHOU Y, LI L, et al. HIF-1α protects against oxidative stress by directly targeting mitochondria[J]. Redox Biology, 2019, 25: 101109. |
| [61] |
GOULDING R P, CHARLTON B T, BREEDVELD E A, et al. Skeletal muscle mitochondrial fragmentation predicts age-associated decline in physical capacity[J]. Aging Cell, 2025, 24(2): e14386. |
| [62] |
HOSGOOD H D R, DAVITT M, CAWTHON R, et al. Mito-chondrial DNA fragmentation and risk of non-Hodgkin lymphoma[J]. JAMA Network Open, 2023, 6(8): e2326885. |
| [63] |
TIAN J, HUANG Y, WU T, et al. The use of Chinese Yang/Qi-invigorating tonic botanical drugs/herbal formulations in ameliorating chronic kidney disease by enhancing mitochondrial function[J]. Frontiers in Pharmacology, 2021, 12: 622498. |
| [64] |
LUO J, SHEN S, XIA J, et al. Mitochondria as the essence of Yang Qi in the human body[J]. Phenomics, 2022, 2(5): 336-348. |
| [65] |
LEONG P K, WONG H S, CHEN J, et al. Yang/Qi invigoration: An herbal therapy for chronic fatigue syndrome with yang deficiency?[J]. Evidence-Based Complementary and Alternative Medicine, 2015, 2015: 945901. |
| [66] |
曹顺金, 张自强. 中医药对阳虚型便秘的治疗进展[J]. 湖南中医杂志, 2020, 36(9): 190-191. |
| [67] |
罗春晖, 蒋宜伟, 安宁宁, 等. 基于"脾与小肠相通"理论谈脾与肠道微生态和骨质疏松症的相关性[J]. 中国微生态学杂志, 2023, 35(5): 599-602. |
| [68] |
中华中医药学会脾胃病分会. 肝脾不调证中医诊疗专家共识(2023)[J]. 中国中西医结合消化杂志, 2024, 32(12): 1033-1039. |
| [69] |
ZHANG Z, YANG S, LIN X, et al. Metabolomics of spleen-Yang deficiency syndrome and the therapeutic effect of Fuzi Lizhong pill on regulating endogenous metabolism[J]. Journal of Ethnopharmacology, 2021, 278: 114281. |
| [70] |
ZHANG S, ZHAO L, HOU X, et al. International clinical practice guideline on the use of traditional Chinese medicine for functional dyspepsia(2025)[J]. Journal of Integrative Medicine, 2025, 23(5): 502-518. |
| [71] |
HE M, LIM X Y, LI J, et al. Mechanisms of acupuncture at Zusanli(ST36) and its combinational acupoints for stress gastric ulcer based on the correlation between Zang-fu and acupoints[J]. Journal of Integrative Medicine, 2025, 23(1): 1-11. |
| [72] |
BAE S, JANG Y, KIM Y, et al. Gut microbiota regulation by acupuncture and moxibustion: A systematic review and meta-analysis[J]. The American Journal of Chinese Medicine, 2024, 52(5): 1245-1273. |
| [73] |
DENG X, LIU J, SUN X, et al. Benzoylaconine induces mitochondrial biogenesis in mice via activating AMPK signaling cascade[J]. Acta Pharmacologica Sinica, 2019, 40(5): 658-665. |
| [74] |
ZHANG D, CHENG H, WU J, et al. The energy metabolism-promoting effect of aconite is associated with gut microbiota and bile acid receptor TGR5-UCP1 signaling[J]. Frontiers in Pharmacology, 2024, 15: 1392385. |
| [75] |
WU S, WANG L, WANG F, et al. Resveratrol improved mitochondrial biogenesis by activating SIRT1/PGC-1α signal pathway in SAP[J]. Scientific Reports, 2024, 14(1): 26216. |
| [76] |
赵罗娜, 杨元凤, 黎豫川, 等. 附子-肉桂调控结肠VIP/cAMP/PKA/AQP通路改善大鼠阳虚型慢传输型便秘[J]. 中国实验方剂学杂志, 2023, 29(21): 103-113. |
| [77] |
JIANG B, YANG Y, DANG W, et al. Astragaloside Ⅳ reverses simvastatin-induced skeletal muscle injury by activating the AMPK-PGC-1α signalling pathway[J]. Phyto-therapy Research, 2020, 34(5): 1175-1184. |
| [78] |
XU M, MA Q, FAN C, et al. Ginsenosides Rb1 and Rg1 protect primary cultured astrocytes against oxygen-glucose deprivation/reoxygenation-induced injury via improving mitochondrial function[J]. International Journal of Molecular Sciences, 2019, 20(23): 6086. |
| [79] |
DONG Y, REN C, TANG H, et al. Icariin promotes skeletal muscle mitochondrial biogenesis by the AMPK-PPARδ-PGC-1α signaling pathway: A potential mechanism for beneficial muscle function homeostasis[J]. Food Bioscience, 2024, 62: 105260. |
| [80] |
CAGGIANO E G, TANIGUCHI C M. UCP2 and pancreatic cancer: Conscious uncoupling for therapeutic effect[J]. Cancer and Metastasis Reviews, 2024, 43(2): 777-794. |
| [81] |
ZHANG R, CAO S, SHI Y, et al. Astragaloside Ⅳ-mediated inhibition of oxidative stress by upregulation of ghrelin in type 2 diabetes-induced cognitive impairment[J]. Naunyn-Schmiedeberg's Archives of Pharmacology, 2023, 396(10): 2637-2650. |
| [82] |
HAN J, SU G, WANG Y, et al. 18β-Glycyrrhetinic acid improves cardiac diastolic function by attenuating intracellular calcium overload[J]. Current Medical Science, 2020, 40(4): 654-661. |
| [83] |
CHOI E M. Glabridin protects osteoblastic MC3T3-E1 cells against antimycin A induced cytotoxicity[J]. Chemico-Biological Interactions, 2011, 193(1): 71-78. |
| [84] |
LI X, LIN Q, GOU F, et al. Effects of hesperidin on mitochondrial function, mitochondria-associated endoplasmic reticulum membranes and IP3R-MCU calcium axis in the intestine of piglets exposed to deoxynivalenol[J]. Food & Function, 2024, 15(12): 6459-6474. |
| [85] |
KAVALENIA T A, LAPSHINA E A, ILYICH T V, et al. Functional activity and morphology of isolated rat cardiac mitochondria under calcium overload: Effect of naringin[J]. Molecular and Cellular Biochemistry, 2024, 479(12): 3329-3340. |
| [86] |
ZHENG B, QI J, YANG Y, et al. Mechanisms of cinnamic aldehyde against myocardial ischemia/hypoxia injury in vivo and in vitro: Involvement of regulating PI3K/AKT signaling pathway[J]. Biomedicine & Pharmacotherapy, 2022, 147: 112674. |
| [87] |
CHOI Y H. Schisandrin A prevents oxidative stress-induced DNA damage and apoptosis by attenuating ROS generation in C2C12 cells[J]. Biomedicine & Pharmacotherapy, 2018, 106: 902-909. |
| [88] |
LI C, JI K B, KIM E Y, et al. Schisandrin B mitigates oxidative damage in early porcine embryos exposed to ochratoxin A via NRF2 pathway activation[J]. Animal Reproduction Science, 2025, 279: 107889. |
| [89] |
ZHENG M, LIU Y, ZHANG G, et al. The antioxidant properties, metabolism, application and mechanism of ferulic acid in medicine, food, cosmetics, livestock and poultry[J]. Antioxidants, 2024, 13(7): 853. |
| [90] |
CAO D, HUANG L, LU X, et al. Potential of ferulic acid as a food-derived cardioprotective agent targeting mitochondrial dysfunction[J]. Food Bioscience, 2025, 73: 107704. |
| [91] |
GUI T, CHEN Q, LI J, et al. Astragaloside Ⅳ alleviates 1-deoxysphinganine-induced mitochondrial dysfunction during the progression of chronic kidney disease through p62-Nrf2 antioxidant pathway[J]. Frontiers in Pharmacology, 2023, 14: 1092475. |
| [92] |
CHU S, ZHANG Z, ZHOU X, et al. Ginsenoside Rg1 protects against ischemic/reperfusion-induced neuronal injury through miR-144/Nrf2/ARE pathway[J]. Acta Pharmacologica Sinica, 2019, 40(1): 13-25. |
| [93] |
KONG L, LIU Y, LI J, et al. Ginsenoside Rg1 alleviates chronic inflammation-induced neuronal ferroptosis and cognitive impairments via regulation of AIM2-Nrf2 signaling pathway[J]. Journal of Ethnopharmacology, 2024, 330: 118205. |
| [94] |
SUI C, LIU Y, JIANG J, et al. Ginsenoside Rg1 ameliorates cerebral ischemia-reperfusion injury by regulating Pink1/Parkin-mediated mitochondrial autophagy and inhibiting microglia NLRP3 activation[J]. Brain Research Bulletin, 2024, 216: 111043. |
| [95] |
ZHU H, YAN C, YAO P, et al. Ginsenoside Rg1 protects cardiac mitochondrial function via targeting GSTP1 to block S-glutathionylation of optic atrophy 1[J]. Free Radical Biology and Medicine, 2023, 204: 54-67. |
| [96] |
ZHANG X, CUI S, ZU Y, et al. Antioxidant properties of acteoside against biological systems: Hemoglobin and cardiomyocyte as potential models[J]. Arabian Journal of Chemistry, 2024, 17(4): 105630. |
| [97] |
ZHANG S, GONG F, LIU J, et al. A novel PHD2 inhibitor acteoside from Cistanche tubulosa induces skeletal muscle mitophagy to improve cancer-related fatigue[J]. Biomedicine & Pharmacotherapy, 2022, 150: 113004. |
| [98] |
CHEN Y, TIAN Y, GUAN B, et al. Morinda officinalis oli-gosaccharides attenuate mitochondria-associated ferroptosis via the NOX4/mitoGPX4 pathway in myocardial ischemia? Reperfusion injury[J]. Frontiers in Cell and Developmental Biology, 2025, 13: 1605513. |
| [99] |
YANG L, AO Y, LI Y, et al. Morinda officinalis oligosac-charides mitigate depression-like behaviors in hypertension rats by regulating Mfn2-mediated mitophagy[J]. Journal of Neuroinflammation, 2023, 20(1): 31. |
| [100] |
ZHAI Y, FENG L, MO Z, et al. 6-Gingerol and 6-shogaol attenuate chemotherapy-induced nausea and vomiting via ameliorating mitochondrial dysfunction and pyroptosis through JNK/MFF signaling pathway[J]. Food Bioscience, 2025, 69: 106808. |
| [101] |
HAN X, YANG Y, ZHANG M, et al. Protective effects of 6-gingerol on cardiotoxicity induced by arsenic trioxide through AMPK/SIRT1/PGC-1α signaling pathway[J]. Frontiers in Pharmacology, 2022, 13: 868393. |
| [102] |
HUANG T, FU H, HO C, et al. Induction of apoptosis by cinnamaldehyde from indigenous cinnamon Cinnamomum osmophloeum Kaneh through reactive oxygen species production, glutathione depletion, and caspase activation in human leukemia K562 cells[J]. Food Chemistry, 2007, 103(2): 434-443. |
| [103] |
WEN S, WANG L, ZHANG C, et al. PINK1/Parkin-mediated mitophagy modulates cadmium-induced apoptosis in rat cerebral cortical neurons[J]. Ecotoxicology and Environmental Safety, 2022, 244: 114052. |
| [104] |
KE S, ZHU W, LAN Z, et al. Cinnamaldehyde regulates mitochondrial quality against hydrogen peroxide induced apoptosis in mouse lung mesenchymal stem cells via the PINK1/Parkin signaling pathway[J]. PeerJ, 2022, 10: e14045. |