 2026, Vol. 45
2026, Vol. 45文章信息
- 曹秀楠, 欧阳慧子, 陈路, 陈涛, 范栢爽, 高英杰, 柴士伟
- CAO Xiunan, OUYANG Huizi, CHEN Lu, CHEN Tao, FAN Baishuang, GAO Yingjie, CHAI Shiwei
- 益肾养肝合剂有效成分抗骨质疏松的机制研究进展
- Research progress on the mechanisms of the active ingredients in Kidney- and Liver-Nourishing Compound against osteoporosis
- 天津中医药大学学报, 2026, 45(6): 746-752
- Journal of Tianjin University of Traditional Chinese Medicine, 2026, 45(6): 746-752
- http://dx.doi.org/10.11656/j.issn.1673-9043.2026.06.15
-
文章历史
收稿日期: 2026-02-20

引用本文 |



2. 中医国家临床医学研究中心,天津 300193;
3. 天津中医药大学,组分中药国家重点实验室,天津 301617
2. National Clinical Research Center for Chinese Medicine, Tianjin 300193, China;
3. State Key Laboratory of Component-based Chinese Medicine, Tianjin University of Traditional Chinese Medicine, Tianjin 301617, China
骨质疏松症(OP)是一种以骨量减少,骨组织微结构损坏,导致骨脆性增加,易发生骨折为特征的全身代谢性骨病。OP作为一种慢性全球性公共卫生问题,在世界范围内患病率逐年增加 [1]。根据最新流行病学研究显示,OP在中国65岁以上人群中的患病率高达46.45%,具有患病率高、治疗费用高、致残率高的特点,严重危害老年人群的生活质量 [2]。目前OP的治疗药物主要为促骨形成剂,如甲状旁腺素类似物;抑制骨吸收剂,如双膦酸盐类;骨矿化相关药物,如维生素D及钙剂 [3]。这些药物存在一定的不良反应如头晕、心悸、高钙血症和药物依赖性等,长期使用可能会限制治疗的安全性 [4-6],故研发防治OP的替代药品及补充药物势在必行。益肾养肝合剂以补气升阳之黄芪为君药,使脾气健旺,中气足,运化正常,化生水谷精微以养先天,辅以山茱萸补肝肾,益精髓以及淫羊藿等补阳之品,共为臣药。其临床使用多年,疗效显著,未发现不良反应。现将近年益肾养肝合剂各有效成分抗OP的骨代谢调节机制研究综述如下,以期为今后的研究和临床用药提供参考。
1 益肾养肝合剂抗OP的化学成分黄芪化学成分复杂,含有多糖类,糖苷类及黄酮类等 [7],其中黄芪多糖和黄芪甲苷为发挥抗OP作用的主要药效成分 [8]。山茱萸主要药效成分为环烯醚萜及其苷、三萜、黄酮等。山茱萸中的莫诺苷 [9]、山茱萸新苷Ⅰ[10]、山茱萸总苷 [11]可促进成骨细胞(OB)的增殖和分化、马钱苷 [12]能抑制破骨细胞(OC)分化和骨吸收、减少骨质流失,从而发挥抗OP作用 [13]。淫羊藿主要含有黄酮类,木脂素类及多糖类等成分,黄酮类化合物是其主要活性成分。通过各种体内外实验结果累积证明,淫羊藿总黄酮具有良好的抗OP疗效 [14],具有促进骨形成和抑制骨吸收的双重活性。益肾养肝合剂抗骨质疏松药效成分化学信息,见表 1。
| 药效成分 | 所属药物 | 化合物类别 | 化学结构 | CAS号 |
| 毛蕊异黄酮葡萄糖苷 | 黄芪 | 黄酮类 | 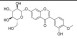
|
20633-67-4 |
| 黄芪多糖 | 黄芪 | 多糖类 | 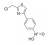 |
89250-26-0 |
| 黄芪甲苷 | 黄芪 | 糖苷类 | 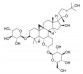 |
83207-58-3 |
| 山茱萸新苷Ⅰ | 山茱萸 | 环烯醚萜苷 | 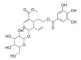 |
131189-57-6 |
| 山茱萸总苷 | 山茱萸 | 糖苷类 | - | - |
| 莫诺苷 | 山茱萸 | 糖苷类 | 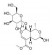 |
25406-64-8 |
| 马钱苷 | 山茱萸 | 环烯醚萜苷 | 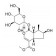 |
18524-94-2 |
| 淫羊藿苷 | 淫羊藿 | 黄酮类 | 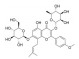 |
489-32-7 |
| 朝藿定A | 淫羊藿 | 黄酮类 | 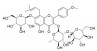 |
110623-72-8 |
| 朝藿定B | 淫羊藿 | 黄酮类 | 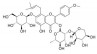 |
110623-73-9 |
| 朝藿定C | 淫羊藿 | 黄酮类 | 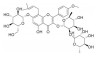 |
110642-44-9 |
| 宝藿苷-Ⅰ | 淫羊藿 | 黄酮类 | 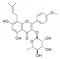 |
113558-15-9 |
骨代谢涉及多类骨细胞及其复杂的信号调控网络,主要包括OB介导的骨形成与OC介导的骨吸收,两者共同维持骨组织的动态平衡。OB和OC通过分泌多种旁分泌因子,实现彼此间的精细调控,这对于骨稳态的维持至关重要。一旦OC数量或活性异常升高,骨吸收速率超过骨形成,骨代谢平衡便会被打破,进而诱发OP等疾病 [15]。近年研究表明,益肾养肝合剂中有效成分可通过多途径促进骨髓间充质干细胞(BMSCs)与OB的增殖分化,同时诱导OC凋亡,从而调节骨代谢、改善骨质疏松。
2.1 体外作用机制 2.1.1 促进BMSCs向OB分化BMSCs来源于中胚层和骨髓内,能够分化形成OB、脂肪细胞、软骨细胞及成肌细胞等 [16-17],其中成骨和成脂肪分化是最主要的两个分化方向。幼年和青壮年时期人的BMSCs主要分化为OB,进入老年后则主要分化为脂肪细胞,从而导致OB数量减少,引起OP的发生。
Jian等 [18]发现毛蕊异黄酮葡萄糖苷可以通过上调骨形态发生蛋白(BMP)/Wnt信号通路 [19]的调节因子β-连环蛋白(β-catenin)、BMP2、p-Smad1/5/8、核心结合因子2(Runx2)的蛋白表达,进而提升成骨标志物碱性磷酸酶(ALP)、促进骨钙素(Ocal)的mRNA表达水平,从而诱导小鼠骨髓基质贴壁ST2细胞增殖分化,促进骨再生。Cai等 [17]通过体外细胞实验发现黄芪甲苷可以介导转化生长因子(TGF)-β/Smad信号通路促进BMSCs的成骨分化。王家伟等 [20]进一步验证上述结果,并明确黄芪甲苷通过作用于TGF-β1/Smad2通路,增加NG2、α-SMA mRNA的表达并提高p-Smad2蛋白水平而发挥作用。翟鑫祥等 [21]发现黄芪多糖可以同时提高去卵巢(OVX)大鼠的BMP2及骨保护素(OPG)蛋白表达水平,促进BMSCs向OB分化。
Yu等 [22]研究发现淫羊藿苷具有双向调节作用,能够促进抗凋亡基因B细胞淋巴瘤因子2(Bcl-2)、拮抗促凋亡基因Bcl-2相关X蛋白(Bax)的表达,从而增加进入S期的细胞数量,显著促进人间充质干细胞的增殖速率并有效抑制其凋亡。Li等 [23]通过实验验证,淫羊藿苷能够促进BMSCs增殖并能下调成脂分化标志物过氧化物酶相关因子的表达,进而促进OB分化,同时抑制脂肪细胞分化。核心结合因子2(Runx2)是诱导BMSCs分化成熟的关键因子,淫羊藿苷可以上调β-catenin和Runx2的蛋白表达水平,促进BMSCs成骨分化,抑制其成脂分化并提高钙沉积面积 [24]。
2.1.2 促进OB增殖、分化OB由BMSCs分化而来,主要负责Ⅰ型骨胶原(Col Ⅰ)等骨基质的合成和分泌以及骨基质的矿化 [25]。OB与新骨生成过程密切相关,因此,促进OB的增殖分化,从而增强骨形成,进而减轻OP是临床常用的治疗手段。
黄芪多糖为黄芪最主要的活性成分,近年来关于黄芪多糖促进OB增殖、分化的机制研究愈趋广泛。研究表明,黄芪多糖可以上调ALP、Runx2以及钙沉积标志物Col Ⅰ蛋白的表达,进而促进OB的增殖、分化,且呈剂量相关 [26]。另外,黄芪多糖能够通过增强BMP2的表达,并提高其下游信号分子细胞外信号调节激酶(ERK)、丝裂原活化蛋白激酶(MAPK)和磷酸化p38丝裂原活化蛋白激酶(P-p38MAPK)的磷酸化水平,从而促进大鼠原代OB的增殖、分化及矿化过程 [27]。柴艺汇等 [28]研究揭示了黄芪多糖能够上调OB维生素D受体mRNA及蛋白的表达,并调节1α-羟化酶(CYP27B)和24-羟基化酶(CYP24A)的mRNA及蛋白表达水平,从而增强OB活性 [29]。孙文星 [30]等发现黄芪多糖通过磷脂酰肌醇3-激酶/蛋白激酶B/哺乳动物雷帕霉素靶蛋白(PI3K/AKT/mTOR)通路诱导细胞自噬,促进OB增殖。
黄佳纯等 [10]研究发现山茱萸新苷Ⅰ能够上调大鼠原代OB中Wnt/β-catenin信号通路和蛋白质二硫键异构酶表达水平,在浓度为1 mmol/mL时能明显增强OB中Wnt2、BMP2、β-catenin、OPG和NADPH氧化酶4(NOX4)的mRNA含量表达水平,使蛋白激酶R样内质网激酶(PERK)的mRNA含量下降;增加Wnt2、β-catenin、蛋白质二硫键异构酶(PDI)的蛋白表达量,降低免疫球蛋白重链结合蛋白(BIP)的表达量,最终促进OB的增殖及成骨分化。山茱萸总苷对防治大鼠OP有明确疗效,主要通过调控骨组织中瞬时受体电位香草酸通道6(TRPV6),钙通道(TRPV5)蛋白表达情况,优化二者倍比关系,改变OB、OC的增殖分化行为,最终提高骨密度而达到防治骨质疏松的疗效 [11]。莫诺苷可通过靶向钠-葡萄糖协同转运蛋白2(SGLT2)促进OB的增殖、分化,改善糖皮质激素诱导的OP[31]。
淫羊藿苷是从淫羊藿中分离的主要活性类黄酮葡萄糖苷,是骨骼中的一种合成代谢剂,已被广泛证明具有抗骨质疏松活性 [32],可促进骨形成和减少骨吸收。淫羊藿苷能够影响雌激素受体介导的ERK和c-Jun N末端激酶(JNK)信号传导途径,促进OB的增殖并抑制细胞凋亡。此外,淫羊藿苷还能提升ALP和Col Ⅰ的表达水平,从而促进骨结节的形成,有效防治OP[33]。
2.1.3 抑制OC活性OC是体内唯一具有骨吸收功能的细胞,通过伪足附着在旧骨区域释放盐酸及组织蛋白酶K(CTSK)降解骨基质,分解骨蛋白行使骨吸收功能。它由骨髓造血干细胞分化而来,在巨噬细胞集落刺激因子(M-CSF)、核因子κB受体活化因子(RANK)、核因子κB受体活化因子配体(RANKL)[34]、OPG的共同刺激下分化成熟 [35-37]。
黄芪多糖能够显著提高骨质疏松大鼠的骨密度,有效改善其骨微结构,降低抗酒石酸酸性磷酸酶(ACP5)和肿瘤坏死因子-α(TNF-α)、白细胞介素-2(IL-2)的含量来抑制OC的分化。在维持OC和OB动态平衡中黄芪多糖发挥了重要作用:它下调了结肠上皮组织FBJ成骨肉瘤癌基因B(Fosb)的表达,使得OC分化通路中活化蛋白1(AP-1)蛋白的产生减少,从而抑制了OC的分化,促进OB的生成;它促进了位于线粒体的雷诺受体(RYR)和位于内质网和肌质网(ER/SR)的G蛋白偶联受体(GPCR)和B细胞受体(BCR)的产生,从而增加结肠上皮组织钙的释放,增强骨组织对钙的利用度,是黄芪多糖改善骨质疏松的一个新的机制 [38]。Wang等 [39]通过实验证明黄芪甲苷可以促进OB的细胞活力、成骨分化和血管生成基因表达,刺激成骨和血管生成的耦合,加速骨再生。
莫诺苷可通过激活PI3K/Akt/mTOR通路促进OB前体转化为成熟的OB[9],并维持骨细胞稳态,同时抑制抗酒石酸酸性磷酸酶(TRAP)的活性、下调活化T细胞核因子1来阻止OC分化 [40]。马钱苷能上调ALP、骨γ-羧基谷氨酸蛋白等OB诱导标志物的表达水平,而下调酸性磷酸酶5、CTSK和基质金属蛋白酶9的表达以抑制OC分化和骨吸收、促进成骨、减少骨质流失,从而有效预防OP[41]。
研究表明,淫羊藿苷能够调节OC内部的微环境,其通过降低细胞内钙离子(Ca2+)水平,显著减少骨吸收陷窝的面积,从而延缓OC介导的骨吸收过程 [42]。Xu等 [43]进一步发现,淫羊藿苷可通过抑制MAPK和核因子κB(NF-κB)信号通路,阻断RANKL诱导的OC分化。此外,该成分还能抑制纤维状肌动蛋白(F-actin)环的生成,降低成熟OC的骨吸收活性 [44]。有研究还指出,淫羊藿苷可通过调控RANKL介导的TNF受体相关因子6(TRAF-6)/NF-κB/ERK信号轴,抑制Raw 264.7细胞向破骨细胞的分化 [45]。
2.2 体内作用机制体内作用机制主要围绕促进骨形成及抑制骨丢失两方面展开。薛志远 [46]研究发现,黄芪煎剂可以提高OVX雌性大鼠全身及右股骨的骨密度,同时增加血清中碱性磷酸酶(AKP)的活性,并降低酸性磷酸酶(StrACP)的活性。黄芪总黄酮提取物能有效提高骨质疏松模型大鼠的骨密度,明显抑制骨矿的溶解和丢失。雌激素具有双向调节的作用,可以在促进OB活性的同时抑制OC的活性,从而加速骨的生长,减少钙质沉积 [47]。徐忠明等 [48]以黄芪为君药的中药复方对绝经后骨质疏松女性进行了干预,结果显示该方剂不仅有效缓解了肾阳虚相关症状,还可能通过提升体内性激素水平、抑制白细胞介素-6(IL-6)介导的骨吸收过度反应,从而发挥骨保护作用。
Xue等 [35]通过蛋白质组学分析以及血清代谢组学分析,对OVX大鼠的股骨进行了深入研究,发现淫羊藿苷不仅能提高骨密度和骨细胞数量,还能改善细胞骨架结构,上调OB中Runx2和成骨细胞特异性转录因子(Osterix)mRNA表达,降低降钙素受体(CTR)、碳酸酐酶Ⅱ(CAⅡ)mRNA表达水平。Cao等 [49]采用OVX大鼠模型,发现淫羊藿苷不仅促进骨痂形成和骨愈合,还能提升骨密度,并有效降低血清Ocal、ALP及TRAP水平,提示其对骨形成和骨吸收均有调控作用。马慧萍等 [50]通过动物实验进一步证实,淫羊藿总黄酮能够通过保护性腺功能、抑制骨吸收及促进骨形成,使骨代谢维持在骨形成大于骨吸收的正向平衡状态,从而减少骨量丢失,延缓骨质疏松进展。其有效成分抗骨质疏松的骨代谢调节机制见表 2和图 1。
| 化学成分 | 作用通路 | 主要分子靶点 | 效应 | 模型 | 参考文献 |
| 山茱萸总苷 | ↑Ca2+通道途径 | TRPV6/TRPV5基因表达倍比关系 | 促进BMSCs向OB分化 | 体外: 大鼠股骨骨组织RNA提取 | [11] |
| 黄芪甲苷 | ↑TGF-β1/Smad2通路 | NG2↑、α-SMA↑、p-Smad2↑ | 促进BMSCs向OB分化 | 体外: rBMSCs | [17, 20] |
| 淫羊藿苷 | ↓信号通路 | Bcl-2↑、Bax↓、JNK↓、c-Jun↓ | 促进BMSCs向OB分化 | 体外: 人间充质干细胞 | [22] |
| 淫羊藿苷 | ↓ER信号通路 | rBMSCs的脂肪分化↓、PPARγ↓、C/EBPα↓ | 促进BMSCs向OB分化 | 体外: rBMSCs | [35] |
| 莫诺苷 | ↑PI3K/AKT/mTOR信号通路 | PI3K↑、AKT↑、mTOR↑ | 促进OB增殖及分化 | 体外: 小鼠OB系MC3T3-E1、小鼠原代培养的OB和OC | [9] |
| 莫诺苷 | ↑SGLT2通道途径 | ALP↑、Runx2↑、Col-Ⅰ↑ | 促进OB增殖及分化 | 体外: OB系MC3T3-E1 | [31] |
| 莫诺苷 | - | ALP↑、Runx2↑、Sp7↑ | 促进OB增殖及分化 | 体外: 小鼠OB系MC3T3-E1 | [40] |
| 山茱萸新苷Ⅰ | ↑Wnt/β-catenin信号通路及ERS途径 | Wnt2↑、BMP2↑、β-catenin↑、OPG↑、NOX4↑、PERK↑、PDI↑、BIP↓ | 促进OB增殖及分化 | 体外: 大鼠原代OB | [10] |
| 毛蕊异黄酮葡萄糖苷 | ↑BMP/Wnt信号通路 | β-catenin↑、BMP2↑、p-Smad1/5/8↑、Runx2↑、ALP↑、Ocal mRNA↑ | 促进OB增殖及分化 | 体外: 小鼠骨髓基质贴壁ST2细胞 | [18] |
| 黄芪多糖 | ↑β-catenin信号通路 | β-catenin↑、ALP↑、Runx2↑、Ocal↑、Col Ⅰ | 促进OB增殖及分化 | 体外: 小鼠OB系MC3T3-E1 | [26] |
| 黄芪多糖 | ↑BMP2、MAPK信号通路 | BMP2↑、ERK MAPK↑、P38 MAPK↑ | 促进OB增殖及分化 | 体外: 大鼠原代OB | [27] |
| 黄芪多糖 | ↑维生素D-FGF-23-Klotho轴 | VDR↑ | 促进OB增殖及分化 | 体外: 小鼠OB系MC3T3-E1 | [28] |
| 黄芪多糖 | ↑维生素D-FGF-23-Klotho轴 | CYP24A↑、CYP27B↑ | 促进OB增殖及分化 | 体外: 小鼠OB系MC3T3-E1 | [29] |
| 黄芪多糖 | ↑PI3K/AKT/mTOR信号通路 | 自噬体↑、Beclin-1↑、P62↓、P-AKT↑、P-mTOR↑ | 促进OB增殖及分化 | 体外: BMSCs诱导分化为OB | [30] |
| 淫羊藿苷 | ↑ERK和JNK信号通路 | Cyclin E↑、PCNA↑; Cdkn2B↓、半胱氨酸天冬氨酸蛋白酶-3活性↓ | 促进OB增殖及分化 | 体外: MC3T3-E1小鼠OB | [33] |
| 淫羊藿苷 | ↑MAPK信号传导、Ca2+信号传导 | Runx2↑、Osterix↑、CTX-Ⅰ↓、CTR↓、CAII↓、P38↓、JNK↓ | 促进OB增殖及分化 | 体内: 雌性ICR小鼠 | [35] |
| 黄芪甲苷 | ↑AKT/GSK-3β/β-catenin通路 | PDGF-BB诱导的血管生成↑ | 抑制OC活性 | 体外: rBMSCs | [39] |
| 淫羊藿苷 | - | Ca2+↓、细胞内超氧阴离子自由基↓ | 抑制OC活性 | 体外: 大耳白兔OC | [42] |
| 淫羊藿苷 | ↓NF-κB和MAPK信号通路 | NF-κB↓、MAPK↓、F-actin↓ | 抑制OC活性 | 体外: 骨髓巨噬细胞BMMs、RAW264.7细胞 | [43] |
| 淫羊藿苷 | ↓TRAF-6/NF-κB/ERK信号通路 | MCF7↓、MDA MB-231↓、TRAF-6表达↓、ERK磷酸化↓ | 抑制OC活性 | 体外: RAW264.7细胞 | [44] |
| 淫羊藿苷 | ↓Rho GTPases和FAK蛋白 | F-actin环↓、FAK↓ | 抑制OC活性 | 体外: OC | [45] |
| 黄芪多糖 | ↑PI3K/AKT/mTOR信号通路 | BMD↑、BS/BV↑、Tb.N↑、Tb.Th↑、Tb.Sp↑ | 促进成骨作用 | 体内: 糖皮质激素诱导的骨质疏松症模型大鼠 | [30] |
| 黄芪多糖 | - | ACP5↓、TNF-α↓、IL-2↓ | 抑制OC活性 | 体内: 大鼠血清 | [38] |
| 淫羊藿苷 | - | 骨痂形成↑、骨愈合↑、骨密度↑、Ocal↓、AKP↓、TRAP↓ | 促进成骨作用 | 体内: 骨质疏松大鼠模型 | [49] |
| 淫羊藿总黄酮 | - | 股骨的干重↑、灰重↑、骨钙↑、骨磷↑、胫骨近端的骨小梁面积百分比↑、骨小梁厚度↑、骨小梁间隙↑、胫骨中段的皮质骨面积百分比↑ | 促进成骨作用 | 体内: 大鼠 | [50] |
| 水煎液 | - | 腰背疼痛缓解时间↓、VAS评分↑; 第4腰椎、股骨颈、桡骨远端骨密度↑ | 促进成骨作用 | 体内: 患者 | [51] |
| 注:PPARγ,过氧化物酶体增殖物激活受体γ;C/EBPα,CCAAT/增强子结合蛋白α;ERS,内质网应激;Cyclin E,细胞周期蛋白E;PCNA,增殖细胞核抗原;Cdkn2B,细胞周期蛋白依赖性激酶抑制因子2B;CTX-Ⅰ,Ⅰ型胶原交联C-末端肽;GSK-3β,糖原合成酶激酶-3β;PDGF-BB,血小板衍生生长因BB;BMD,骨密度;BS/BV,骨表面积/骨体积;Tb.N,骨小梁数量;Tb.Th,骨小梁厚度;Tb.Sp,骨小梁分离度。 | |||||

|
| 图 1 益肾养肝合剂有效成分通过调控骨代谢抗骨质疏松的机制图 |
OP的防治是医学领域面临的难题,随着药理学、分子生物学的快速发展,应用中药防治多发、疑难性疾病的研究日渐兴盛。中国步入老龄化社会、OP发病率日益上升,中药复方治疗OP效果显著,优势突出,其有效成分和作用机制已经成为研究热点。通过总结可以看出,益肾养肝合剂中的药效成分(毛蕊异黄酮葡萄糖苷、黄芪多糖、黄芪甲苷、山茱萸新苷Ⅰ、山茱萸总苷、莫诺苷、马钱苷、淫羊藿苷、淫羊藿总黄酮)通过调衡骨稳态干预OP的机制可分为促进BMSCs向OB分化,促进OB增殖、分化,抑制OC活性,其中涉及了OPG/RANK/RANKL、Wnt/β-catenin、PI3K/AKT/mTOR、BMP/Smad、ERK、JNK等多种信号通路及细胞因子。随着该领域研究的不断深入,有利于学者全面认识OP发病机制并为开发有效的治疗药物提供科学依据。
| [1] |
CHENG C, WENTWORTH K, SHOBACK D M. New Frontiers in Osteoporosis Therapy[J]. Annual Review of Medicine, 2020, 71: 277-288. DOI:10.1146/annurev-med-052218-020620 |
| [2] |
黄宏兴, 晁爱军, 程群, 等. 医疗机构骨质疏松专科建设专家共识[J]. 中国骨质疏松杂志, 2024, 30(6): 781-789. |
| [3] |
LANE N E. Epidemiology, etiology, and diagnosis of osteoporosis[J]. American Journal of Obstetrics and Gynecology, 2006, 194(2): S3-S11. DOI:10.1016/j.ajog.2005.08.047 |
| [4] |
Cosman F, Langdahl B, Leder B Z. Treatment Sequence for Osteoporosis[J]. Endocrine Practice, 2024, 30(5): 490-496. DOI:10.1016/j.eprac.2024.01.014 |
| [5] |
SRIVASTAVA M, DEAL C. Osteoporosis in the elderly: prevention and treatment[J]. Clinics in Geriatric Medicine, 2002, 18(3): 529-555. DOI:10.1016/S0749-0690(02)00022-8 |
| [6] |
PENG Z, XU R, YOU Q. Role of Traditional Chinese Medicine in Bone Regeneration and Osteoporosis[J]. Frontiers in Bioengineering and Biotechnology, 2022, 10: 911326. DOI:10.3389/fbioe.2022.911326 |
| [7] |
LI X, QU L, DONG Y, et al. A review of recent research progress on the astragalus genus[J]. Molecules(Basel, Switzerland), 2014, 19(11): 18850-18880. DOI:10.3390/molecules191118850 |
| [8] |
刘路, 李凯, 胡阳, 等. 黄芪有效成分抗骨质疏松症作用机制的研究进展[J]. 中草药, 2023, 54(4): 1321-1328. |
| [9] |
LIU H, LI X, LIN J, et al. Morroniside promotes the osteogenesis by activating PI3K/Akt/mTOR signaling[J]. Bioscience, Biotechnology, and Biochemistry, 2021, 85(2): 332-339. DOI:10.1093/bbb/zbaa010 |
| [10] |
黄佳纯, 林燕平, 陈桐莹, 等. 山茱萸新苷Ⅰ对成骨细胞的增殖及成骨分化的影响[J]. 中国骨质疏松杂志, 2020, 26(1): 1-5. |
| [11] |
李绍烁, 赵京涛, 何昌强, 等. 山茱萸总甙干预骨质疏松模型大鼠骨代谢: TRPV6、TRPV5通路的变化[J]. 中国组织工程研究, 2019, 23(11): 1749-1754. |
| [12] |
LU Y, LUO Q, JIA X, et al. Multidisciplinary strategies to enhance therapeutic effects of flavonoids from Epimedii Folium: Integration of herbal medicine, enzyme engineering, and nanotechnology[J]. Journal of Pharmaceutical Analysis, 2023, 13(3): 239-254. DOI:10.1016/j.jpha.2022.12.001 |
| [13] |
TANG X, HUANG Y, FANG X, et al. Cornus officinalis: A potential herb for treatment of osteoporosis[J]. Frontiers in Medicine, 2023, 10: 1289144. DOI:10.3389/fmed.2023.1289144 |
| [14] |
HUANG M, XU S, LIU L, et al. m6A Methylation Regulates Osteoblastic Differentiation and Bone Remodeling[J]. Frontiers in Cell and Developmental Biology, 2021, 9: 783322. DOI:10.3389/fcell.2021.783322 |
| [15] |
HE X, HU W, ZHANG Y, et al. Cellular senescence in skeletal disease: mechanisms and treatment[J]. Cellular and Molecular Biology Letters, 2023, 28(1): 88. DOI:10.1186/s11658-023-00501-5 |
| [16] |
UZIELIENE I, BERNOTIENE E, RAKAUSKIENE G, et al. The Antihypertensive Drug Nifedipine Modulates the Metabolism of Chondrocytes and Human Bone Marrow-Derived Mesenchymal Stem Cells[J]. Frontiers in Endocrinology, 2019, 10: 756. DOI:10.3389/fendo.2019.00756 |
| [17] |
CAI B, ZHANG A G, ZHANG X, et al. Promoting Effects on Proliferation and Chondrogenic Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells by Four "Kidney-Tonifying" Traditional Chinese Herbs[J]. BioMed Research International, 2015, 792161. |
| [18] |
JIAN J, SUN L, CHENG X, et al. Calycosin-7-O-β-d-glucopyranoside stimulates osteoblast differentiation through regulating the BMP/WNT signaling pathways[J]. Acta Pharmaceutica Sinica B, 2015, 5(5): 454-460. DOI:10.1016/j.apsb.2015.06.005 |
| [19] |
GAO Y, CHEN N, FU Z, et al. Progress of Wnt Signaling Pathway in Osteoporosis[J]. Biomolecules, 2023, 13(3): 483. DOI:10.3390/biom13030483 |
| [20] |
王家伟, 孙廉旭, 张莎, 等. 黄芪甲苷激活TGF-β1/Smad2信号通路诱导骨髓间充质干细胞向周细胞分化的作用研究[J]. 现代生物医学进展, 2018, 18(8): 1407-1413. |
| [21] |
翟鑫祥, 董辉, 王晶, 等. 黄芪多糖保护OVX大鼠成骨细胞功能活性的机制研究[J]. 天津医药, 2022, 50(3): 265-269. |
| [22] |
YU T, XIONG Y, LUU S, et al. The shared KEGG pathways between icariin-targeted genes and osteoporosis[J]. Aging, 2020, 12(9): 8191-8201. DOI:10.18632/aging.103133 |
| [23] |
LI X, PENG B, PAN Y, et al. Icariin stimulates osteogenic differentiation and suppresses adipogenic differentiation of rBMSCs via estrogen receptor signaling[J]. Molecular Medi-cine Reports, 2018, 18(3): 3483-3489. |
| [24] |
涂艳, 熊莉娜, 柳湘洁, 等. 淫羊藿苷对成骨细胞成骨分化的影响及Wnt/β-catenin信号系统的关系研究[J]. 中国中医急症, 2017, 26(3): 448-450, 466. |
| [25] |
VULF M, KHLUSOV I, YUROVA K, et al. MicroRNA Regulation of Bone Marrow Mesenchymal Stem Cells in the Development of Osteoporosis in Obesity[J]. Frontiers in Bioscience(Scholar Edition), 2022, 14(3): 17. DOI:10.31083/j.fbs1403017 |
| [26] |
胡俊, 杨盼盼, 袁忠, 等. 黄芪多糖对MC3T3-E1细胞增殖及成骨分化的影响[J]. 中国骨质疏松杂志, 2021, 27(12): 1788-1791, 1799. |
| [27] |
孔祥鹤, 牛银波, 王婷梅, 等. 黄芪多糖对大鼠原代成骨细胞的影响及其机制研究[J]. 中草药, 2011, 42(10): 2065-2069. |
| [28] |
柴艺汇, 高洁, 陈云志, 等. 黄芪多糖对小鼠MC-3T3-E1成骨细胞维生素D受体mRNA及蛋白表达的影响[J]. 时珍国医国药, 2018, 29(6): 1281-1283. |
| [29] |
柴艺汇, 高洁, 田兴中, 等. 黄芪多糖对MC-3T3-E1成骨细胞CYP27B, CYP24A mRNA及蛋白表达的影响[J]. 中国实验方剂学杂志, 2018, 24(13): 147-151. |
| [30] |
孙文星, 黄万新, 刘传慧, 等. 黄芪多糖通过PI3K/AKT/mTOR促进激素性骨质疏松症大鼠成骨细胞增殖[J]. 中国骨质疏松杂志, 2023, 29(1): 35-40. |
| [31] |
YANG H Z, DONG R, JIA Y, et al. Morroniside ameliorates glucocorticoid-induced osteoporosis and promotes osteoblastogenesis by interacting with sodium-glucose co-transporter 2[J]. Pharmaceutical Biology, 2023, 61(1): 416-426. DOI:10.1080/13880209.2023.2173787 |
| [32] |
DONG H, TANG F, ZHAO Z, et al. The Bioactive Compounds of Epimedium and Their Potential Mechanism of Action in Treating Osteoporosis: A Network Pharmacology and Experimental Validation Study[J]. Pharmaceuticals, 2024, 17(6): 706. DOI:10.3390/ph17060706 |
| [33] |
SONG L, ZHAO J, ZHANG X, et al. Icariin induces osteoblast proliferation, differentiation and mineralization through estrogen receptor-mediated ERK and JNK signal activation[J]. European Journal of Pharmacology, 2013, 714(1-3): 15-22. DOI:10.1016/j.ejphar.2013.05.039 |
| [34] |
ZHU M, GUO Q, KANG H, et al. Inhibition of FAAH suppresses RANKL-induced osteoclastogenesis and attenuates ovariectomy-induced bone loss partially through repressing the IL17 pathway[J]. FASEB Journal, 2023, 37(1): e22690. DOI:10.1096/fj.202200911R |
| [35] |
XUE L, JIANG Y, HAN T, et al. Comparative proteomic and metabolomic analysis reveals the antiosteoporotic molecular mechanism of icariin from Epimedium brevicornu maxim[J]. Journal of Ethnopharmacology, 2016, 192: 370-381. DOI:10.1016/j.jep.2016.07.037 |
| [36] |
KITAURA H, MARAHLEH A, OHORI F, et al. Osteocyte-Related Cytokines Regulate Osteoclast Formation and Bone Resorption[J]. International Journal of Molecular Sciences, 2020, 21(14): 5169. DOI:10.3390/ijms21145169 |
| [37] |
WANG C, LI F, LI Y, et al. Cistanche Deserticola for Regulation of Bone Metabolism: Therapeutic Potential and Molecular Mechanisms on Postmenopausal Osteoporosis[J]. Chinese Journal of Integrative Medicine, 2023, 29(1): 74-80. DOI:10.1007/s11655-022-3518-z |
| [38] |
刘军. 基于组学研究黄芪多糖改善大鼠骨质疏松的机制[D]. 广州: 暨南大学, 2021.
|
| [39] |
WANG F, QIAN H, KONG L, et al. Accelerated Bone Re-generation by Astragaloside Ⅳ through Stimulating the Coupling of Osteogenesis and Angiogenesis[J]. International Journal of Biological Sciences, 2021, 17(7): 1821-1836. DOI:10.7150/ijbs.57681 |
| [40] |
LEE C G, KIM J, YUN S H, et al. Anti-Osteoporotic Effect of Morroniside on Osteoblast and Osteoclast Differentiation in Vitro and Ovariectomized Mice in Vivo[J]. International Journal of Molecular Sciences, 2021, 22(19): 10642. DOI:10.3390/ijms221910642 |
| [41] |
LEE C G, KIM D W, KIM J, et al. Effects of Loganin on Bone Formation and Resorption in vitro and in vivo[J]. International Journal of Molecular Sciences, 2022, 23(22): 14128. DOI:10.3390/ijms232214128 |
| [42] |
黄健, 张金超, 张天蓝, 等. 淫羊藿苷对破骨细胞骨吸收功能的影响及其作用机制[J]. 科学通报, 2006, 57(24): 2851-2856. |
| [43] |
XU Q, CHEN G, LIU X, et al. Icariin inhibits RANKL-induced osteoclastogenesis via modulation of the NF-κB andMAPK signaling pathways[J]. Biochemical and Biophysical Research Communications, 2019, 508(3): 902-906. DOI:10.1016/j.bbrc.2018.11.201 |
| [44] |
郭元晖, 薛黎明, 聂燕, 等. 淫羊藿苷和仙茅苷协同抑制破骨细胞的形成、分化和骨吸收功能[J]. 药学实践杂志, 2013, 31(4): 262-266. |
| [45] |
KIM B, LEE K Y, PARK B. Icariin abrogates osteoclast formation through the regulation of the RANKL-mediated TRAF6/NF-κB/ERK signaling pathway in Raw264.7 cells[J]. Phytomedicine, 2018, 51: 181-190. DOI:10.1016/j.phymed.2018.06.020 |
| [46] |
薛志远. 红芪和黄芪抗骨质疏松、抗氧化及抗肝损伤作用比较研究[D]. 兰州: 兰州大学, 2018.
|
| [47] |
QIAO S, ZHANG X, CHEN Z, et al. Alloferon-1 ameliorates estrogen deficiency-induced osteoporosis through dampening the NLRP3/caspase-1/IL-1β/IL-18 signaling pathway[J]. International Immunopharmacology, 2023, 124: 110954. DOI:10.1016/j.intimp.2023.110954 |
| [48] |
徐忠明, 周志昆. 黄芪三仙汤治疗绝经后骨质疏松症36例临床研究[J]. 中医药导报, 2009, 15(5): 9-11. |
| [49] |
CAO H, ZHANG Y, QIAN W, et al. Effect of icariin on fracture healing in an ovariectomized rat model of osteoporosis[J]. Experimental and Therapeutic Medicine, 2017, 13(5): 2399-2404. DOI:10.3892/etm.2017.4233 |
| [50] |
马慧萍, 贾正平, 葛欣, 等. 淫羊藿总黄酮抗大鼠实验性骨质疏松作用研究[J]. 华西药学杂志, 2002, 17(3): 163-167. |